OK, so let me make some points to begin with.
What I did was, I implemented the Mendelian Laws Of Inheritance into a PureData patch that sends out MIDI data to controlle synthesizers (hardware as well as software). It’s like a genetic MIDI controller, so to speak.
Synthesizers have a limited potential of generating sounds, so they span a finite search space for my MIDI controller to operate on.
The genetic algorithm is setting the synthesizer’s sound manipulating parameters to random values, first. And this 8 times for one population of sounds.
It’s kind of like Richard Dawkins’ “Biomorphs” program - only with sounds instead of graphics.
Biomorphs are phenotypic representations of certain combinations of parameter values, as you can see here:
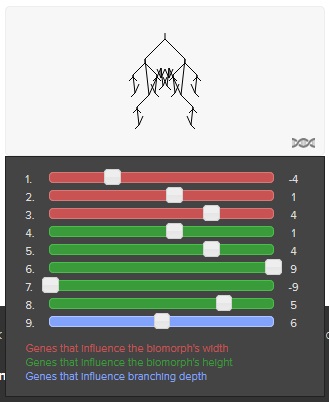
All sliders controlle certain visual aspects of the graphical structure of Biomorphs.
8 random Biomophs have 8 random sets of parameter values for their 9 parametric genes.
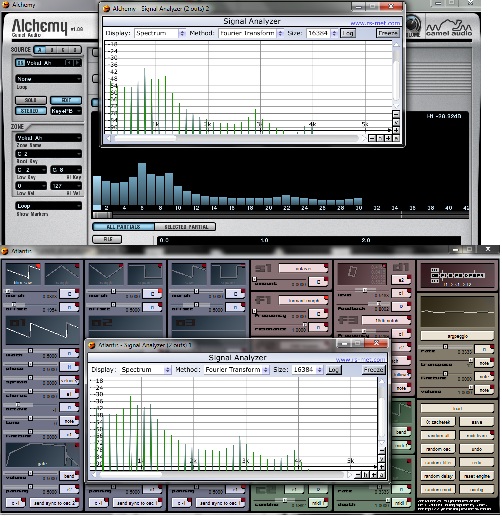
Synthesizers depict parameters in the same way, as you can see here:

So it’s obviously possible to use a genetic algorithm to controlle synths the same way like Biomorphs - and that’s exactly what I did.
The genetic algorithm is now suggesting a couple of phenotypes to the user, out of which he can now select those of interest and breed further by combining the earlier selected (parent) sounds with new random sounds - generating offspring sounds. Some parameters are inherited from that parent sounds and some are inherited from a selected mating partner sound. The offspring sound is therefore a certain combination of both parent sound and mating partner sound.
The way the parameter values are inherited is either by dominant-recessive inheritance or intermediate inheritance. Dominant-recessively inherited parameter values stay unchanged - either they come from parent sound or mating partner sound. Intermediately inherited parameter values are an interpolation between the respective values of the parent or the mating partner sound.
The offspring sound is a complex combination result of both.
Now, what’s obvious is, that no new parameters are generated. There won’t be any new parametric genes generated by the algorithm. The synthesizer stays the same, of course, but by recombination and selection alone, the algorithm forces the sounds to adapt to the user’s selection criteria.
As the synthesizers can of course be exchanged, different synthesizers will have differently formed search spaces for the algorithm to look for sounds in.
If I now set the search criterion to the most brilliant periodic sound, the algorithm will soon find sounds that are pretty comparable - no matter what synthesis method I use.
See the following spectra:
Different synthesis methods - same sounds. All without new parameters generated. Only the parameter values of each synthesizer have been changed.
This shows: Different genotypes can generate same or at least similar phenotypes without adding new information to the synthesizer’s source codes.
Hence, you don’t need mutations to explain phenotypic similarity.
All it takes is recombination of already available parameters in parameterized systems.
Now, I am convinced that life forms are equally parameterized systems.
All sexually reproducing life forms adapt according to the Mendelian Laws Of Inheritance in connection with epigenetic reactions to the environment.
The a-sexually reproducing life forms vary due to epigenetics alone.
The question arising from that insight is, if mutations are really needed for any kind of empirically observable adaptation.
My answer would be, No.
The reason why I think so, is, that the fundamental precondition for evolution to be possible is a continuity from species to spiecies - without any limits. The evolutionist perspective is, that mutations are the driving force of evolution, as far as I know, right?
But how could one actually prove that claim?
What my algorithm shows is, that adaptation can be caused by recombination alone in finite parameter search spaces. So, if there were no mutations at all, for argument’s sake, animals could only adapt within certain limits - by recombination and epigenetics alone.
To evidence evolution, though, you’d have to find an animal phenotype that would not have been genetically encoded within these limits. But if you don’t know where exactly these limits are; how will you be able to determine a phenotype that’s positioned >outside< of these limits?
You cannot possibly tell! Why? Because variation is not only caused by recombination of monogenetic traits, but also by polygenetic traits and epigenetic reactions to the environment - expanding the limits into the unknown. Additional to that, genetically hereditable traits are molecular-biologically not even clearly definable, so where do you draw the line between phenotypes encoded within the limits set by recombination and epigenetics and phenotypes beyond that limits? And how do you determine where the variation range of one taxonomic family ends and where that of another animial’s taxonomic family begins?
Say you found an animal that you definately know to be not encoded in the variation range of a certain taxonimic family. What if it is part of another animal’s taxonomic family that’s neighboring it?
You could not possibly tell if any animal phenotype is the result of recombination + epigenetics or mutation! Therefore, you cannot prove a continuity of species.
Hence, the major precondition for evolution is - well… not scientifically provable. It’s just an assumption taken by belief.
I hope, I could express myself understandable, anyhow…
Regards.