I thought this merited a conversation of its own. I’m posting the question here, and the first answer by @deliberateresult.
Continuing the discussion from Irreducible complexity and mere complexity:
I thought this merited a conversation of its own. I’m posting the question here, and the first answer by @deliberateresult.
Continuing the discussion from Irreducible complexity and mere complexity:
So I think we still stuck on a very abstract definition:
But what is, for example, the body plan difference between humans and chimps? Do we not have the same body plan?
And how did the evolution of wings coming into this? I thought the whole point of body plans was to acknowledge that wings do NOT require a new body plan, but share common history (or design) with limbs.
Can someone from outside the theistic evolutionist camp (or at least in the ID camp) try and explain how you think about this? Thanks!
I do object. This is false, and you are misrepresenting evolutionary theory. Every step does not need to convey a selective advantage. Who exactly are you arguing with? Just a straw man strict-Darwinist?
@deliberateresult Thanks for your response! I think it is possible to ignore drift as you suggest, if we are not too detailed and focus on the broad outlines of evolution, because while small changes do occur due to drift, an overall change in body shape and function should have some adaptive purpose.
I am afraid you will have to wait a longish time for a detailed genotypic narrative of any evolution from long before DNA remains could have been preserved. It is possible scientists may eventually reconstruct such a narrative from existing bird (and crocodile) DNA, but in the meantime, let’s assume you are not just seeking questions beyond the current frontiers of science. Science will always have frontiers.
I think we will have better luck with a phenotypic step by step functional adaptive continuum. Where do we start? “No wings.” Okay. But the bones in the (bird) wing have been known for ages to correspond to the forelimb of mammals, lizards, and other tetrapods. So let’s start with that.
The dinosaurs most closely related to birds all appear to have been bipedal, and cursorial (runners on two legs). Indeed, the ancestor of all dinosaurs was bipedal and cursorial. So what was the functional advantage of picking the forelimbs up off the ground and not using them?
There can be a number of different advantages to bipedalism, but to pick just one likely culprit, modern-day lizards will sometimes run on their back legs in order to sprint. Sprinting is useful in hunting, and it fits the description of Euraptor as a small hunter which could sprint very fast.
So, what to do with the forearms now? In some lineages, for example famous T. Rex, the forearms weren’t used for much except perhaps grabbing prey, and became very small, nearly vestigial. But in some lineages, like Maniraptora, they developed long forearms with three long fingers.
Note that feathers had already been evolving for some time by this point, as many non-flying dinosaurs are known to have had feathers: for warmth, display, etc.
A lot of people have argued whether birds evolved flight from trees, like flying squirrels, or from running and jumping on the ground. Trees seem intuitively likely, but there’s little evidence of tree-climbing adaptations in the fossils. One suggestion from 2004 is that they hunted by leaping and pouncing on prey from above, which would encourage the gradual improvement of gliding and aerodynamic form, without requiring true flight capability, but cheerfully selecting for it when it did eventually evolve. I like this hypothesis for several reasons. It’s easy to picture a sprinting predator shifting to ambush-style hunting, it provides a clear driver for selective change, and it occupies a middle ground in the tree-climbing/ground-running debate.
I think I’ve covered each functionally adaptive step. If you would like to flesh out details, it would be very useful to look up all the dinosaurs considered most closely related to birds; Microraptor is an interesting one, and so is Anchiornis. Arcteryx is the famous one, of course, but it already has bird-like wings, so it’s necessary to go back further if we want to look for the evolution of the wing.
Flight evolved independently several times: in birds, in bats, and in pterosaurs (like pterodactyls); in insects like beetles, and we even see intermediate forms of flight (which fall short of true flight) in flying fish and flying squirrels. I didn’t take your question to mean you’d like descriptions of each of these evolutionary pathways (bats, for example, are known to have been tree-climbing in their initial stages) but you are welcome to inquire further!
My point, of course, is that although comparing a modern-day bird to a modern-day lizard looks like an obvious case of a novel body plan, when we go back and look at the fossils, we find lots of intermediates which make it much harder to point to a particular form which sprang up without reference to very similar forms predating it.
I look forward to continuing this discussion! And thanks, @benkirk for your response in the original thread, as well!
I think you hit on exactly some of the questions I’ve been wondering, thank you and thanks for starting a new topic!
The body plan of a whale is a perfect candidate for a discussion like this. Toothed whales have a very specific adjustment to their nasal passage ways.
One might imagine that the left and right nostrils of a spermacetti whale [I intentionally do not use the shorter name for this whale… because it tends to get emails blocked in most academic settings!] proceed right to the blowhole - - at the TOP of its head! - - where a mighty exhale sends the spray high into the air. But no!
The LEFT NASAL PASSAGE has anatomically drifted directly to the ‘vestibulum’ of the blowhole, where the might spray originates.
The RIGHT nasal passage, is even longer … ultimately merging with the LEFT just before the blow hole. But the Sperm whale represents the “compromise”. Read this discussion of alternate “nasal plans”!:
QUOTE:
"In Mysticetes [technical term for the 15 species of baleen whales] the nasal passages are separate tubes all the way to the external nares [nares = nostrils], whereas in Odontocetes [term for 73 species of toothed whales], the two nasal passages branch into a complex series of nasal sacs that eventually coalesce into a single blowhole. The sperm whale is however intermediate as it exhibits a sigmoidally-shaped blowhole formed by two nasal tubes that remain distinct from the bony nares to the top of the head. . . "
So… this is certainly a NOVEL BODY plan, yes?.. in terms of internal anatomical arrangements?
I dunno @gbrooks, is this really a novel body plan? Everything seems homologous to prior body plans…I do not think this is what it means…
HA! I expected an objection! Do you know the real reason I chose whales?
So that no matter what somebody said… there was SOMETHING we could use…
Forget the nasal passages… how about the LACK of rear legs (setting aside for the time being the whales that have vestiges of rear legs) ? Surely having 2 limbs instead of 4 qualifies, yes?
So @Lynn_Munter, doing some research on this, I think “novel body” plans is commonly thought to refer to the explosion of complexity we see in the the Cambrian period the “Cambrian Explosion”. See for example…
http://www.americanscientist.org/issues/pub/the-origin-of-animal-body-plans
The Cambrian Explosion and the “sudden” arrival of “complex” body plans (like those of insects and fish and reptiles rather than just jelly fish, sponges, etc.) here seems entirely surprising from some understandings of evolution. This turns out to be a point that many old earth creationists (like Hugh Ross) point to as evidence against evolution.
So to clarify, even in these arguments, wings are not new body plans. Humans are not new body plans from chimps. Whales are not new body plans either. We all share the same body plans, The question being raised: where did the first body plans come from? And why did they all suddenly appear? @deliberateresult seems to be taking this a different direction, but his point seems to be somewhat idiosyncratic. Though I am not sure. Perhaps he can clarify.
Now, there appear to be several problems with this line of reasoning, not the least of which is that it relies on a misunderstanding of evolution. Evolution is just “common descent” by largely natural mechanisms. It has not consistently made predictions about how quickly new things arise.
That being said, the “Cambrian Explosion” did not actually show the “sudden” arrival of complex body plans. First, there are precursors to these body plans in the age before (Palaeozoic) that show they are not totally novel. Moreover, the Cambrian Explosion period spans about 30 to 40 MILLION years, over which this long period we see these plans arise and diversify. 30 to 40 million years hardly qualifies as “sudden,” by any reasonable definition.
Knowing what we know of genetics now, some of this makes sense. It appears that some parts of the evolutionary pathway (like complex body plans) are harder to evolve than others. However, once that step occurs, it opens up a rapid diversification that elaborates that “innovation” by adapting easily into new functions. The first step is hard and slow (and perhaps dependent on oxygen). The second steps are easier and quicker. The first step is a “hill” evolution has to get over, after which a lot can ‘quickly’ happen in the downhill roll.
I think most modern biologists would hold that complex body plans did not arise as suddenly as we first thought. Nonetheless, this was probably one of those difficult “humps” that took a long time to get over, but once we did it enabled a lot of rapid evolution as the basic innovation was tweaked into different niches.
This article from our friends (with whom we respectfully disagree) at Reasons to Believe provides the best description of a body plan I’ve found as is relevant to us. (my comments are in brackets).
This portion of the biblical creation story bears a remarkable resemblance to an event called the Cambrian explosion. Some 543 million years ago, the number of Earth’s animal phyla (a phylum designates life-forms sharing the same basic body plan)
Cambrian Explosion Brings Burst of Evidence for Creation - Reasons to Believe
Though if you look at phylum, a lot of “body plans” by the definition just look like variations on worms (which by the way predate the Cambrian period). Phylum - Wikipedia I think his next definition is a bit more precise…
Of the 182 animal skeletal designs theoretically permitted by the laws of physics, 146 appear in the Cambrian explosion fossils.
While evolutionary scenarios, as opposed to worked-out theories, exist for hypothesizing how new genera, new orders, and new families of animal life might appear, there is no rational evolutionary scenario for explaining how a new animal phylum might appear. [each phyla they say has a different body plan]
The Cambrian explosion marks the first appearance of animals with skeletons, bilateral symmetry, appendages, brains, eyes, and digestive tracts that include mouths and anuses. [some of this turns out to be outdated; e.g. worms already existed, and without skeletons how do you reliably see brains? So there is some "ascertainment bias here due to the difficulty in fossilizing creatures without skeletons]
The moment oxygen levels in Earth’s atmosphere and oceans permit the existence of Cambrian animals, they suddenly appear. [over 30 million years is not “sudden”, this also points to the mechanism why they did not arise earlier]
The Cambrian explosion includes the most advanced of the animal phyla, chordates, including vertebrate chordates.
Hi Joshua - The comments in brackets [like this] are yours and are not in the original, correct?
No, it’s a modification of the body plan we share with whales. The nostrils on embryonic whales are in the same position as ours.
Correct. Look at the links for yourself.
I think that body plans are at or near the heart of the Gould vs. Conway Morris debate about evolution’s predictability. Gould interpreted the Precambrian fossil record as depicting a plethora of body plans lost forever. Conway Morris (the world’s expert on the topic) sees them very differently, essentially as modifications of existing body plans. That’s a necessarily brief overview of the debate, worthy of discussion elsewhere, but the point is that the deep history of body plans can take us to very interesting considerations of evolutionary constraint (or lack thereof). Gould was arguing that evolution left vast tracts of morphospace unexplored, by burning bridges along a herky-jerky path that could have gone just about any other way. Conway Morris wonders if the opposite is the case, that body plans are almost hard-wired into the fabric of biology.
In any case, I think that when evolutionary biologists and embryologists talk of “body plans” they mean deep and fundamental organizational themes that characterize large and ancient sets of lineages. New nasal turbinates are as far as one can get from that. Thanks for a cool topic!
I suppose I take a Conway Morris approach then.
I should make one revision to my larger point. I think the Cambrian explosion actually is longer than 40 million years, probably closer to 80 million. Perhaps a geologist can help make this more precise? (paging @Joel_Duff, @davidson, and @RyanBebej)
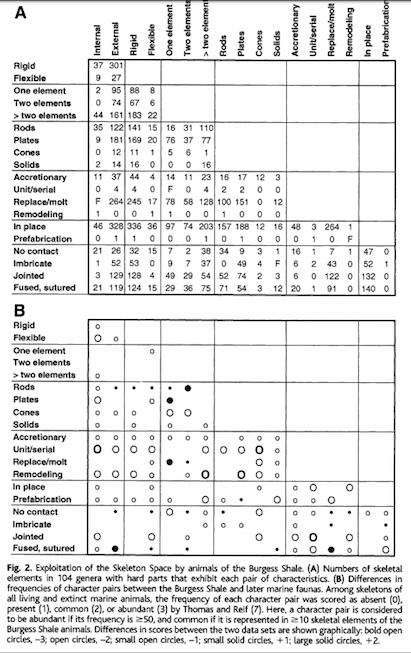
That quote hit my attention too. It references a really fascinating 2008 paper in Science…
The set of viable design elements available for animals to use in building
skeletons has been fully exploited. Analysis of animal skeletons in relation to
the multivariate, theoretical “Skeleton Space” has shown that a large proportion
of these options are used in each phylum. Here, we show that structural
elements deployed in the skeletons of Burgess Shale animals (Middle Cambrian)
incorporate 146 of 182 character pairs defined in this morphospace. Within 15
million years of the appearance of crown groups of phyla with substantial hard
parts, at least 80 percent of skeletal design elements recognized among living
and extinct marine metazoans were exploited.
http://faculty.jsd.claremont.edu/dmcfarlane/bio145mcfarlane/PDFs/cambrian%20designs.pdf
One of the reason I love dealing with this debate is the real gems the conversation continually uncovers. Beautiful paper.
Oh yes, cool, I remember that paper now, I think I discovered it by reading Conway Morris’ Life’s Solution. I tend to side with Conway Morris on predictability, mostly because of convergence. But even the analysis of “skeleton space” must necessarily ignore other ways of building organisms (sponges? cephalopods? plants?). I wonder if we still know whether evolution burned some bridges on the way to the Precambrian. It seems to me that this is likely in principle.
I certainly agree that conversations like this provide opportunities to rummage through the attic for treasures! Oh, BTW, I don’t know you and have only read a few of your posts here, but thanks for defending science! I look forward to seeing more of your thoughts and your work.
Nice to meet you too @sfmatheson. Looking forward to getting you know you better. Thanks for linking to your blog too.
I’ve already found the topic of my first post or two: a paper from last month that identifies a single mutation in the human genome that may explain (at least in part) the dramatic expansion of the cerebral cortex that occurred in our lineage
You might be curious to know that we already covered that article! Evolution by splicing . You should post a link to your article there when it is done.
Now returning to the discussion at hand about “body plans.” I noticed some strange irregularities here. Part of it is a misreading of Ross, and the other part what appears to be a mistake in his part. To recap…
Hugh Ross defines body plan as a phyla. First…
a phylum designates life-forms sharing the same basic body plan
Cambrian Explosion Brings Burst of Evidence for Creation - Reasons to Believe
And then he writes in another article…
The Cambrian explosion refers to the sudden, simultaneous appearance of most of the animal phyla (body plans) that occurred 542–543 million years ago.
…
http://www.reasons.org/blogs/todays-new-reason-to-believe/the-cambrian-explosion-and-evolutionists’-responses
But then goes on to make this statement…
Of the 182 animal skeletal designs theoretically permitted by the laws of physics, 146 appear in the Cambrian explosion fossils.
http://www.reasons.org/blogs/todays-new-reason-to-believe/the-cambrian-explosion-and-evolutionists’-responses
He does not reference this claim, but the only candidate article with those two numbers (128 and 146) is the Science article with the abstract…
The set of viable design elements available for animals to use in building
skeletons has been fully exploited. Analysis of animal skeletons in relation to
the multivariate, theoretical “Skeleton Space” has shown that a large proportion
of these options are used in each phylum. Here, we show that structural
elements deployed in the skeletons of Burgess Shale animals (Middle Cambrian)
incorporate 146 of 182 character pairs defined in this morphospace. Within 15
million years of the appearance of crown groups of phyla with substantial hard
parts, at least 80 percent of skeletal design elements recognized among living
and extinct marine metazoans were exploited.
http://faculty.jsd.claremont.edu/dmcfarlane/bio145mcfarlane/PDFs/cambrian%20designs.pdf
Now the first point I now recognize is that I misread Hugh the first time around. He is not saying that skeleton designs are body plans. That is not the case at all. That is something different entirely, in fact the point of the article is that within each phyla a large range of different skeletal designs can be found.
Second, it seems false to reference the “skeletal designs theoretically permitted by the laws of physics.” There is absolutely nothing in this article about the laws of physics limiting the number of designs to 182, and this appears to be Hugh’s invention.
You can call me Stephen, or maybe Steve after we’ve met at a poster session or something. ![]()
I was afraid I’d been scooped by BL! But there’s plenty more to say about the paper. I’ll post a link here if it doesn’t seem like shameless promotion. I’m writing about the 2015 paper now, then will explain the December paper. I think the story is a an example of overnight birth of a new gene, which should be interesting to everyone. (Except normal people, I admit.)
I don’t read RTB because of its fondness for “invention” as you call it. But I do think that skeleton space is a reasonable approximation of “body plan.” It won’t work for organisms with no skeleton, of course, but it’s a really good start.
Feel free to start your own dedicated thread. Shameless promotion of science is encouraged. Maybe it will balance out some of the shameless promotion of other things that goes on here. ![]() (Just click on the +new topic button at the top of the forum home page https://discourse.biologos.org/ )
(Just click on the +new topic button at the top of the forum home page https://discourse.biologos.org/ )
Ross is reading the article by Thomas, et al. (2000) incorrectly. The authors state that there are 182 possible pairs of body plan features. However, there are many other combinations: singletons, triplets, quadruplets, etc. So I wrote a little Python script to calculate the number of possible body plans based on Thomas’ taxonomy:
import numpy as np
from itertools import combinations
def multiply_elements(a):
val = 1
for e in a:
val = val * e
return val
features = np.array([2,2,3,4,4,2,3])
plans = 0
for i in range(1,8):
combos = combinations(features, i)
for c in combos:
plans += multiply_elements(c)
print("number of possible body plans =", plans)
# "number of possible body plans = 10799"
10,799 is a lot more than 182! Since there are only about 3 dozen animal phyla in biological history, it would seem that less than 1% of the possible body plans have been explored.
In addition, I’m not sure that a phylum necessarily equates to a body plan as described by ID theorists. For example, the phylum Chordata includes these two species:


Eptatretus stouti (Pacific hagfish) and Homo sapiens sapiens (human)
“Let your conversation be always full of grace, seasoned with salt, so that you may know how to answer everyone.” -Colossians 4:6
This is a place for gracious dialogue about science and faith. Please read our FAQ/Guidelines before posting.