“There were a number of lineages leading from primates to humans.” So wrote a Harvard professor of biology and mathematics on Edge.org back in 2005, apparently as already proven but without a citation. Perhaps he didn’t mean that several nonhuman primates each evolved into humans, at least two directly from nonhuman primates into the same fully modern human sex, but perhaps he did. I think there’s something to that possibility but I’m an outsider to evolutionary studies and I couldn’t find another mention of this line of thought when I Googled. Citations: Nowak, Martin (no title); Edge.org (2005), as accessed April 23, 2016; and Brockman, John, ed., What We Believe but Cannot Prove (Harper Perennial, 1st ed. 2006), p. 206. I thought to contact you when I saw http://biologos.org/blogs/dennis-venema-letters-to-the-duchess/understanding-evolution-mitochondrial-eve-y-chromosome-adam (2014), as accessed the same day, and found the comments section on that page apparently closed. I doubt that those two humans we might as well call (even without religious connotations) “Adam” and “Eve” ever met or were of the same generation, because if their lifetimes overlapped by even a fraction of a minute then something must have pushed specifically primate-to-human evolution and then that push must have disappeared just afterwards, or we would have brand-new primate-to-human evolution every century or so and occurring in many places each century. It’s likelier that “Adam” and “Eve” each had reproductive intercourse with a primate who was very similar to themself until evolution occurred again. Due to further evolution among all relevant species, such intercourse would now no longer be reproductive. But I’m still curious: Did evolution from primate into humanity occur more than once for either sex?
Hi Nick, welcome to the Forum! Thanks for your question.
There seems to be some confusion in your writing about the term lineage and about the mechanisms of evolution. I will try to clarify things a bit more.
A lineage describes a path of direct descent through any family. For example, there is a “backward” tree consisting of you, your father and mother, their parents, their parents’ parents, et cetera. Your own descendants would be a “forward” tree. Any line through this tree is a “lineage”.
The current picture is that the population from which humanity evolved consisted of about 10,000 people / hominids. If we descended from a population of about 10,000, it means that you can always find multiple separate lineages through the family tree. However, this does not mean that the evolution from primate to human occurred multiple times separately (i.e., in separate populations). I will now list a few important points that are useful to keep in mind when thinking about evolution:
-
Evolutionary mechanisms such as natural selection operate on the population level, not on individuals.
Of course, changes in individuals can eventually spread to the whole population. But such spreading occurs through mechanisms which act on the entire population. Also, the reproductive unit of humanity is man and woman. So you cannot separate men from women on the population level (the level of analysis for evolution). The differences between men and women are coupled to each other, i.e., they only function together. -
Evolution occurs in any population with selective pressure.
The pressure results in a push to find local solutions. There are moments when populations change very rapidly due to evolution, but evolutionary mechanisms are acting on populations all the time. So it’s better not to speak of evolution “occurring” on specific moments, but rather of changes occurring in populations. -
Because evolutionary mechanisms act on the population level, they can exploit rare occurrences and complex contingencies.
Such contingencies are intimately dependent on the circumstances of the population and the environment. So just the fact that something happened in evolutionary history does not say anything about the likelihood that this will happen again. Currently, it appears that humanity evolved from a single population and hence exploited some specific conditions at the time.
However when we see something occurring multiple times in separate populations, things become more interesting. That’s what we call convergent evolution. Pablo de Felipe wrote an interesting BioLogos article called “Evolution on Purpose: The Inevitability of Intelligent Life?”. I suppose that touches upon the original intention of your question. Here’s a quote from that article:
Once life starts moving on, according to Conway Morris, the forces of natural laws would push and constrain it to paths across a biological landscape that could reach, not unexpectedly, the development of an intelligent being that would occupy a mental niche (of course, that does not mean that these beings had to be necessarily Homo sapiens!). As Conway Morris’ ideas have developed in recent years, he prefers the view of evolution as a search engine that explores the opportunities of life and, rather than just emerging in a pre-existent mental world, discovers it after navigating a constrained “hyperspace” of biological possibilities. We could be the first to set foot on this new beach, but it does not mean that we are entirely unique. As he remarks, the intelligence of other runners in this race of discovery, like corvids, dolphins and great apes, “suggests that equivalent search engines are only a few million years behind ourselves.”
So while evolution can lead multiple times to similar solutions, this has not happened (yet?) for our own unique capacities. I hope this will help you to get a clearer picture of the concepts of evolutionary theory and the evolution of mankind.
You write: “Were there multiple lineages from primate to human?”
While it is easy to imagine multiple offshoots from a common ancestor … how would multiple “lineages” ever MERGE TOGETHER to create a single lineage?
If by lineages meaning distinct species… by definition, it is usually accepted that genuinely different species cannot mate and have fertile offspring.
If by lineages, you mean something akin to:
breeding Collies with Poodles and Dachsunds … as you can see, you are treating “lineages” as “breeds” of a single species.
Hi Casper -
This is a wonderfully clear and lucid explanation of evolution. Thanks!
I’m wondering if the article that prompted this thread might have had in mind the multiple hominid lineages present in the human genome. For example, non-African human populations have about 4% Neanderthal DNA, and a more limited set of populations has 1-2% Denisovan DNA. These are the result of interbreeding between homo sapiens sapiens and the other hominids tens of thousands of years ago.
I have tried to follow this reasoning and have also attempted (without great success I fear) to understand the maths behind the modelling - to begin with, a “bottle neck” of some 10,000 units are postulated, and I gather these have reached some sort of equilibrium over an unspecified period. variations are assumed to have taken place - but with such a number of individuals, does the model require constant interactions, leading to breeding patters, or are these nomadic, in which case an equilibrated model would in all probability fail (the units are scattered over a large area , leading to low concentrations and low probabilities of any interactions). I note that this modelling does not seek a sudden mutation that produces new male and female - so a gradual change from an equilibrated sample of a limited number of units, at low concentrations, makes this modelling appear problematical.
Perhaps you may provide details that would clarify these concerns purely from a modeling perspective - if so I would thank you in advance.
Did you read Dennis Venema’s series on this topic? I think his explanations are very accessible. This is the first part:
I’m not completely on top of the matter because I’m not trained specifically in genetics, but I’ll try to explain it according to my understanding derived from readings of Dennis Venema. I do have experience with genetic algorithms though, but that’s a different part of the story.
I think the argument is mainly derived from information theory. Today, there is a certain amount of genetic diversity within mankind. Based on that, you can place constraints on the minimum original size of the population, using the rate of reproduction. You even don’t need to make assumptions about the mutation rates for that. Dennis explained (in this post) that the genetic variation in human populations is far too large for having resulted from a single ancestral pair in the past 200,000 years or so.
The assumption of ideal mixing is actually very conservative. Assuming restraints on the mixing (as in nomadic populations) would result in even less variation and therefore it would push back the limit even further in time. This would make the likelihood of a single ancestral pair even smaller.
I have tried to follow the reasoning based on genetic diversity, but as this is not my field, I have tried to understand the modelling which seems to me to underpin such a hypothesis. The question of a single pair/couple is irrelevant to this discussion. The main point is to account for the present population of humans, using various models. One model that seems to be in favour is that of a bottleneck, and subsequent variations that are thought to produce the human species, and the current diversity. Other models have accounted for the current size and diversity of human population, so my question is to try and understand the assumptions and mathematics used in the various models.
My major objection on basing it all on genetic diversity is that the thinking appears circular, and thus does not have sufficient self-correction in these models - I may have missed something, but from what I have read, this seems to be the case.
Can you list a model which does not assume a bottle-neck of approximately 10,000 individuals and is able to account just as well for the current amount and distribution of genetic diversity among humanity?
I don’t see the circularity, can you be more explicit about it? Genetic diversity is an observation that stands on itself. Researchers also check how genetic diversity differs between different sub-populations. For example, they have used this to reconstruct a migratory pattern of different populations. We know that the differences between African sub-populations is much larger than the European and Asian populations. This has given support to the migration hypothesis (i.e., that mankind originated in Africa). That hypothesis is again supported by anthropological evidence on basis of archeological findings, et cetera.
Maybe the issue is more that you reject a certain part of the corollary conclusions related to a 10,000 bottleneck. If that is the case, could you describe to me what your objections are?
I have a number of references that deal with the subject, and if you feel it is necessary, I will spend some time compiling them for you. My point however, is to understand the thinking and assumptions behind the various models. To this end, I provide two examples that illustrate this point.
In “Modelling the recent common ancestry of all living humans.” By Douglas L. T. Rohde, Steve Olson, Joseph T. Chang, NATURE | VOL 431 | 30 SEPTEMBER 2004 |, we find: “Here we show that recent common ancestors also emerge from two models incorporating substantial population substructure. One model, designed for simplicity and theoretical insight, yields explicit mathematical results through a probabilistic analysis. A more elaborate second model, designed to capture historical population dynamics in a more realistic way, is analysed computationally through Monte Carlo simulations. These analyses suggest that the genealogies of all living humans overlap in remarkable ways in the recent past. In particular, the MRCA of all present-day humans lived just a few thousand years ago in these models. Moreover, among all individuals living more than just a few thousand years earlier than the MRCA, each present day human has exactly the same set of genealogical ancestors.”
In “How wide and how deep is the divide between population genetics and developmental evolution?” by WAGNER, Biology and Philosophy (2005), a book review, we have “Developmental evolution has resuscitated research questions which were abandoned early in the 20th century when Neo-Darwinian theory of evolution took over as the dominant form of evolutionary theory. But still, the distrust between Neo-Darwinian evolutionary biologists, as population geneticists and developmental biologists is still as real as it was when John Maynard-Smith was concerned about my mental state.”
I add a reference that seems to be more in line with your statements, simply to illustrate the diversity that I detect in this subject matters: In Human population dispersal ‘‘Out of Africa’’ estimated from linkage disequilibrium and allele frequencies of SNPs , by McEvoy et al Genome Research, 21:821–829 2011. “Genetic and fossil evidence supports a single, recent (<200,000 yr) origin of modern Homo sapiens in Africa, followed by later population divergence and dispersal across the globe (the ‘‘Out of Africa’’ model). However, there is less agreement on the exact nature of this migration event and dispersal of populations relative to one another. We use the empirically observed genetic correlation structure (or linkage disequilibrium) between 242,000 genome-wide single nucleotide polymorphisms (SNPs) in 17 global populations to reconstruct two key parameters of human evolution: effective population size (Ne) and population divergence times.”
I think these examples make my point regarding models on this area. My comments concerning equilibrium and interactions between a small number of units dispersed over a large area is my feeling regarding models in general – this approach is rationalised by invoking very lengthy periods out of necessity, and even then I wonder how one may invoke a bottleneck as a starting point (as an assumption in such a model).
This paper “Modelling the recent common ancestry of all living humans” actually does not contradict the idea of a genetic bottleneck. They define the most recent common ancestor as follows:
“If a common ancestor of all living humans is defined as an individual who is a genealogical ancestor of all present-day people, …”
Under this definition, the most recent common ancestor lived within a larger population of humans. It does not say anything about the evolution of mankind or the kind of population from which mankind arose.
That paper shows one modelling approach can deal with the population size and diversity using stochastic techniques and available data on births, deaths, migrations etc., - this approach requires a relatively short period (6-10,000 years). Such a result is contrary to other approaches that require >200,000 years, along with a bottleneck. The most recent common ancestor stochastic model simply shows the population can be modelled by starting with one male and one female and does not require any other assumption.
I have read the entire article and that is not the claim of these authors. They are not aiming to reproduce observed patterns of genetic diversity starting from a single ancestral pair. Rather, they are working backward to figure out how long we have to travel backwards in time to find a person among other people who is the great-great-great-(…)-great-grandfather or -mother of everyone currently alive. Again, this does not say anything about the population size or genetic bottlenecks.
You seem to have misunderstood the definition of a “most recent common ancestor” that these authors are using. It does not mean that the whole human population depends on a single ancestral pair. It just means that all people have this ancestral pair in common. There are still large parts where these family trees do not completely overlap.
Their model concluded that there was some person living on Earth among other people about 6-10,000 years ago who is a direct relative of everyone on Earth today. But that common ancestor was living among other people. This can also be seen clearly from this quote from their paper:
(…) Nevertheless,
our results suggest that the most recent common ancestor for the
world’s current population lived in the relatively recent past—
perhaps within the last few thousand years. And a few thousand
years before that, although we have received genetic material in
markedly different proportions from the people alive at the time,
the ancestors of everyone on the Earth today were exactly the same.
So if we take a random person alive today, and go back 6-10,000 years backwards into his family tree, we will have a huge group of people. Among those people, we will be able to find a single person that is present in everyone’s family tree alive today. If we go back in time another few thousand years, we will again have a group of people. But this time, everyone of those ancestors has a familial connection with everyone alive today. Note that they are talking about a whole population and about a common ancestor living in a population.
There is no contradiction between the results of these authors and the approaches that require >200,000 years to arrive at a single ancestral pair parenting the whole of humanity.
I think you may misunderstand the point I am making, which is to show there are various models that may account for the current population and diversity, and some of these models require relatively short periods, while others long periods. I cannot see why you focus on a non-existent argument related to ancestry, or the need for a most common ancestor (MCA). The point of all of these models is to show descent from some ancestry - so it is obvious that a starting point (MCA) would be part of a model. The questions I am asking are focussed on a need, or lack of such a need, of a bottleneck, to enable such modelling to work. Your other points/arguments are not relevant to the model in question. You may wish to bring your own interpretation for the modelling - but for this discussion, that is beside the point.
My point was that you cannot make such points based on this article. Maybe you have other examples than this? Because this article does not actually prove your point.
The article you have referenced does not present a model that accounts differently for the data on genetics. It does not claim that the current genetic diversity needed only a few thousand years to arise. The only thing these author tried to model is to find a family member common to everyone.
When you understand that the model presented by these authors is completely independent from the genetic bottleneck idea, then you will also understand that these models don’t contradict each other.
If you wish, take another look at the assumptions in this article. These authors made assumptions about migration flows in the current world (I copied the map shown below from the article). They already assume stable migration, stable towns, stable routes, multiple populations.
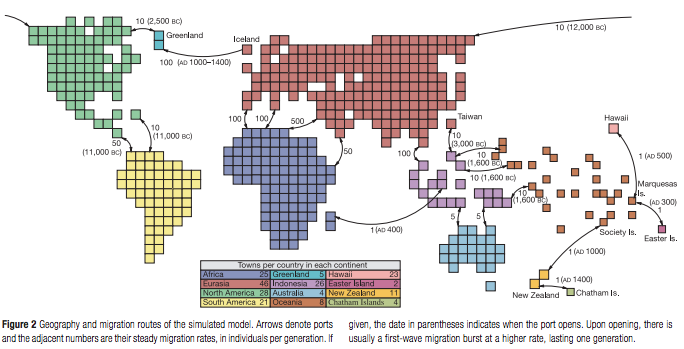
This model does not contradict the need for a genetic bottleneck in any way. The genetic bottleneck took place long before there were so many large migration flows in the world. The key to your question is to realize that these models are actually complementary to each other instead of being different accounts. See this Nature article from 2011 for an insightful review of the evidence for such a bottleneck in human history: Inference of human population history from individual whole-genome sequences | Nature
BTW, the idea of a genetic bottleneck does not in any way presuppose Neo-Darwinism.
There’s really no need to get all cranky about it. You asked me a question. I’m trying to think with you. I asked you for some references. Based on what you have presented, there is no such ambiguity in models as you describe.
My whole premise is that this statement of yours cannot be based on the article that you have provided. That article does not present any alternative model for explaining genetic diversity…
Models can only be compared with each other when they actually aim to reproduce the same phenomenon. But the time periods of these models are actually answering two completely different questions.
-
Given the known migration flows, how long ago was our most recent common family member living on Earth? About 6,000 - 10,000 years.
-
If all of humanity descended from a single ancestral pair (without genetic input from other people), how long ago would that first couple have lived? >200,000 years ago (i.e., too long ago to be consistent with other estimates)
You’re comparing completely different models that try to estimate different kinds of time periods and then stating that their outcomes are variable. This is comparing apples with pears.
Anyway, this is my two cents. Based on what you have presented until now, I don’t see any reason to doubt that there’s a strong consensus within the scientific community regarding the overall outline of the population history of mankind. If you’re not interested in discussing this topic anymore, then we’ll just agree to disagree here. Fine with me ![]() .
.
Hi GJDS,
Having read through this thread, I’d like to comment that Casper has things correct, and his comments to your interpretation of things are on the mark.
You seem to be confusing the concept of finding a common ancestor in the recent past with the idea that all humans descend only from that common ancestor. The paper that discusses a most recent common ancestor (in the last few thousand years) is not saying that all human genetic diversity comes from that most recent common ancestor.
One example of this that I have discussed in detail here is the case of mitochondrial and Y chromosome most recent common ancestors in humans. Yes, we all descend from these individuals, but we also descend from many thousands more. These ideas are not in conflict.
So too with these studies - we may well have a (very) recent common ancestor (for a small part of our DNA), but when looking at the genomes of present day humans as a whole, we see that we descend from a population of ~10,000.
Casper, kudos to you for grasping what the key issues are even though this is not your area of expertise.
Hi @GJDS,
I am sorry that this conversation has turned out to be frustrating for you, it was surely not my intention for it to be so. Let me try one more time to answer your questions according to my understanding.
In this thread, we have encountered two types of models, one focusing on MRCA and one focusing on population genetics. These two models are different in multiple ways, namely in [1] the type of data they use, [2] the kinds of auxiliary assumptions they make and [3] the type of estimate they aim to produce.
MRCA:
[1] population sizes, geography, migration flows (present and historical)
[2] population substructure, mixing
[3] how long ago our most recent common ancestor lived
Population genetics:
[1] distribution of genetic diversity across sub-populations
[2] generation time and (for some methods) mutation rate
[3] minimum initial population size N (or how much time for N=2)
Now, for population genetics, the results [3] are very robust under different methods (different input [1] and different auxiliary assumptions [2] are consistent with initial population size >10,000). There are methods based on linkage disquilibrium (such as the one outlined here) which assume certain mutation rates (also measured, present day). These converge with methods that don’t depend on mutation rates such as coalescent theory. These methods provide lower limits on the population size called the “effective population size”. In case of a lack of interactions between sub-populations (such as with nomadic populations), the population size estimates would become larger because it would effectively reduce the total amount of genetic diversity for a given population size. So the minimum of N>10,000 will always hold. This means that ideal mixing within the population is actually an extremely conservative assumption.
So there are two points. Firstly, I have tried to show that the MRCA model and population genetics models aim to produce completely different kinds of estimates. I’m sorry if that was not the kind of explanation you were looking for, but it does explain why their outcomes don’t match up. Secondly, different approaches to population genetics all independently confirm the conclusion that human population size has never been smaller than 10,000 even under the most conservative assumptions possible. This means that these models are not in anyway circular. Any other reasonable set of assumptions will only act to increase the minimum (so we can’t go any lower than 10,000).
I hope this can help to clear things up.
@DennisVenema, thanks for affirming my conclusions before. If I have made any incorrect statements on the genetics part in this post, please don’t hesitate to correct me.
Hi Casper,
Yes it is regrettable that at times we both may have spoken past each other, and thank you for your patience and time to consider the nature of the models in question.
I will continue to read up further on the techniques used in such studies, partly out of curiosity, but mainly to get a better understanding of the outlook that questions my/our understanding of the account of Adam and Eve.
As I have mentioned in other posts, I do not involve myself in debates dealing with the work of scientists in the bio-areas, but take an interest in terms of gaining insights related to a theoretical framework for biology, as this should form a part of an overall understanding of the sciences. This takes effort and time, and I am not proposing any great change from my efforts, simply a hope to reach a deeper understanding of the sciences in toto.
Perhaps to end this conversation on a constructive, or a useful point, and just out of interest, I make these two (very general) points related to the outcomes from any population model:
(1) based on a general view that we may begin with a small number (relatively) of units, one modelling approach shows that a large section of the current population may have descended from a small number (even N=2). Thus many in Europe may be descendants of Darwin, or a Roman Emperor, and so on. What is the scientific value of such models, in your opinion, and,
(2) those models that consider genetic diversity and mutation often appear (to me at least) to discuss the formation of a human species from pre-, or non-humans. This suggests that mutation rates are related to transformational aspects of descent, and this is central to the arguments for common descent for primates. If this is so, these models (or the modelling approach) should have a predictive capability that shows when another major change may occur on a large time scale.
So purely out of curiosity, can you point to any modelling results in this area that may postulate the next major transformation of humans, or indeed, primates?
Peace and goodwill.
I’m the original poster. Thank you for all of this depth.
I analogized from a claim that eyes evolved into existence at 40 points on an evolutionary tree, which I assume meant that separate origins had been identified. The possibility of multiple human lineages is analogous, but likely limited to one, not 40, because eyes are far simpler than entire primates or humans (who merely include eyes). That “one”, however, does not deny that there could have been an original 10,000 or so, if they were 10,000 or so beings who could reproduce (to yield fertile offspring) with both ancestors and contemporaries who were exclusively primate and contemporaries and descendants who were exclusively human. That would be consistent with a single origin on an evolutionary tree.
I separated women and men in order to solve a problem. Either (a) there’s an in-between state whereby a being (from a potentially small number) can reproduce with either an exclusively primate or an exclusively human or (b) both men and women humans had to come into existence at the same time, i.e., be born within a few years and in sufficiently close proximity so they could reproduce with each other if they couldn’t reproduce with predecessor primates. The former (“(a)”) seems much more likely. If there was an in-between state, there’d still be a first being who occupied that state and every single one of whose contemporaries had to be primates, and that first being could be of either sex. Another being in that same in-between state and who was of the other sex could have been born after the first had died because the the first, in this model or notion, could have reproduced with either primates or humans, as long as there were members in that in-between state from when the first of either sex was born until the first of the other sex was born and able to reproduce. If there was a break in that membership before the appearance of the second sex, the arrival of the second sex would have meant that evolution from primate to the in-between state occurred twice, and I think you’re saying there’s no evidence of that, so the likelihood is of continuity and thus that the firsts of both sexes need never have met, reproduction having been possible without those two having the same offspring (as in, “Eve” mating with some male of some species and having a human child and “Adam” mating with some female of some species and having a human child).
I see that modern humans reproduced both with Neandertals and with Denisovans (https://www.sciencedaily.com/releases/2016/03/160328133514.htm, as accessed May 5, 2016, citing an article in Current Biology). I wasn’t thinking of that when I posted above, as I didn’t know about it except maybe as a recent brief hint on radio, and the original quotation was from 2005, so that author probably didn’t think of it, either. But if that lateral reproduction was possible, then reproduction between earlier and later through an in-between state should be less disputable.
On whether there was an Adam and an Eve 4.3 million years ago regardless of names and theology, I think it impossible that there was not. Even if the DNA of all 10,000 or so plus of 10,000 immediate ancestors was preserved today and analyzed and we were nonetheless unable to determine a dividing line for either sex between clearly primate and ambivalently post-primate, even if bodies’ soft tissue had been likewise preserved and nonetheless we were today unable to find the dividing line visually, there still had to be a first of each sex. There could have been more than one for either sex or even for each sex, but that’s unlikely (and I think you agree), because that would mean multiple occurrences of evolution from primate to human within a single sex and why there would have been two then but none any other time would need explaining. But once per sex makes sense, thus “Adam” and “Eve” existed, even if they never were alive at the same time or in geographic proximity. That makes the in-between state likely to have lasted through many generations.
I’m counting humanity as having started roughly 4.3 million years ago, quite apart from all of us today tracing our own ancestries to two individuals living only about 200,000 years ago. I don’t think that anyone’s claiming that our human ancestors of 200,000 years ago were themselves parented by primates, merely that other humans of 200,000 years ago were not our own principal ancestors (maybe the others contributed bits we can still find but without being principal contributors of our genetic make-up today).
My notion of an in-between state would support findings of diversity and is consistent with a population size of 10,000 or so while still allowing for a first male and a first female who may never have met (I’m assuming no ability back then for an intermediary to preserve and carry live sperm, thus proximity in time and place and thus a meeting would have been necessary for reproduction but only if reproduction was directly between “Adam” and “Eve”, and it likely was not direct).
Nomads may not have had less reproductive contact with non-nomads. Perhaps they did, especially if that’s been empirically found. But I’m not sure nomads had to travel with much less per-square-mile density than non-nomads lived. And, in traveling, they may have passed multiple non-nomadic communities. Socially, nomads could have engaged in reproductive contact with non-nomads more easily because, not living with the non-nomads, they’d be gone and have fewer consequences to worry about. Also, some nomadic groups could have traveled roughly in parallel, such as if they were following the same large herds of animals for food but were otherwise mostly separate, and groups traveling in parallel might reproduce together.
Possibly off-point, a claim I just read is that development of life and of humanity was paced such that Earth’s age could not have been less than it is for us to still be at the stage of development we now have. That would mean that most of life, including humans, have evolved as fast, or nearly as fast, as could have been.
@gbrooks9: Multiple lineages of one species could reproduce with each other, but, in addition to that and in addition to the dog-breed model, we could consider the in-between state. It seems unlikely that the DNA or chromosome of an individual has an identifier that says “I’m a human and no one else can reproduce with me” or “I’m a primate and . . .”. Thus, as evolution proceeds to evolve a new species, there’s an in-between state in which fertile offspring can be produced with beings on either side of the in-between state. Imagine a manufacturer of padlocks that are designed to be opened with different keys but in which two of the padlocks have keys that are so similar that sometimes the key from one opens the other lock, because the engineering tolerances for the keys were just loose enough to unintentionally allow that possibility.
@GJDS: As an outsider to this discipline, regarding the future, I’ll speculate that the timing of evolution from either primate or human to another species will depend on the constraint on a current species’s ability to survive through reproduction. If external conditions, such as predation against a species or toxicity in its diet, become tougher but not so rapidly that evolution can’t offer a solution, then a new species may replace the one that dies off. Without that pressure, evolution can still of course occur and the resulting species can survive to reproduce, but its population would long be outnumbered by the ancestral species and, with a lack of a clear division so that temporally near generations will likely be able to reproduce with each others’ services, it’s more likely that the new would-be species would not become a species but would be absorbed (or its genetically-driven characteristics would be absorbed) into the ancestral species, and we may not be able to tell that that happened to any particular individuals. I don’t know how many primates there are (there are over 300 species (https://nationalzoo.si.edu/Animals/Primates/Facts/, as accessed May 6, 2016)), but, with 7,000,000,000+ humans occupying, I think, most of the Earth’s surface except polar regions and apparently doing biologically well, I doubt that such pressure is likely to arise against humans anytime soon, or against primates, who occupy much less geography but still a lot, some separated by oceans. I’m not sure what you mean by “transformational aspects of descent” but my guess is that the only models that could predict when the next complex species will appear would be models about the world’s changes affecting the survival of a given species, family, or other taxonomic unit or of all species, not a model about the species itself. While I think random mutation occurs at a steady rate, whether the life form resulting from a given mutation survives is due to internal and external conditions, and probably no model can predict survival based on internal conditions for random mutations that are unspecified in advance. That leaves predictability as limited to external conditions, requiring highly complex models of the world. I’m not sure we have the computational power to analyze that (compare to the need for supercomputers just to predict the weather and weather is only one element of a world model for species survival).