Most scientists agree that the origins of life required the ability to produce energy and to store heritable information. The large protein molecules necessary for life’s many functions are constructed by transcription from DNA into mRNA and then transcribed into proteins. But these very DNA functions require many large protein molecules for their activities. Thus it is reasonable for a committed atheist who studies this to begin to believe that perhaps there is a God who began it all and then turned it loose for us to evolve ad lib. And to remain honest to himself, he might restrict his beliefs to that exact point in time and nothing afterward.
So if a God was necessary to create the system necessary for all life, no matter how early or small, whether bacteria, plant or animal, what are the implications? Was He necessary to be involved after that? Could evolution have simply taken the course that we now understand and see? After all, without the Permian extinction, dinosaurs would not have evolved, and without the K-T extinction mammals would not have flourished. Natural events give structure to Earth today, but did it need a nudge at the origin of life? I am only posing these questions as possibilities for an atheist to begin his conversion with the “small step” of a belief in God.
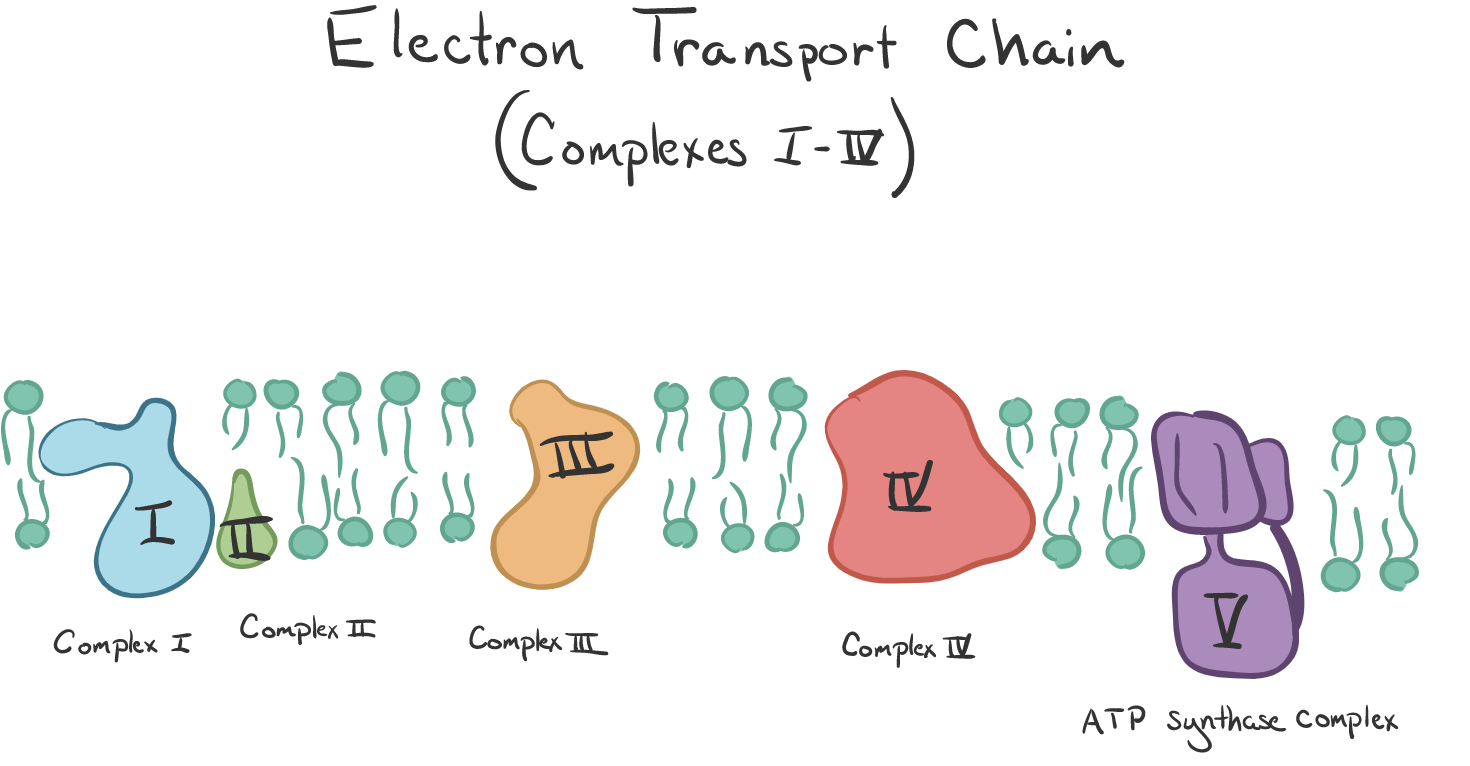
One of these mysteries and possibilities surrounds the common denominator of all energy production in every living cell. This sine-qua-non of energy is the system in mitochondria called the Electron Transport Chain (ETC). Using laser induced liquid bead ion desorption (LILBID) mass spectrometry, we now know the actual structure of the Complexes I-IV that strip and transport electrons and protons (H+). And the final complex, complex V is the one known as ATP synthase. So here is what that scientist/convert to God might conclude:
- Then it may not be necessary to view Christ’s birth as from a virgin.
After all, the Christian idea of a virgin birth is derived from the verse in Isaiah 7:14 describing an “alma” as giving birth. The word “alma” has always meant a young woman, but Christian theologians came centuries later and translated it as “virgin.”
-
The resurrection of Christ is not necessary. Accepting resurrection of the dead is almost impossible to any biologist anyway.
-
All of the miracles are no longer necessarily factual. That life’s events do not follow the natural laws of physics and biology has always been a barrier.
-
The concept of the power of prayer is rendered moot. Of course, a divine God with the power to structure DNA for “origins” could do most anything, but this converted scientist might be comforted that accepting this is no longer necessary.
-
The concept of heaven and hell is discarded. After all, it was difficult to imagine in the first place.
So what name is given to a belief system that accepts a divine origin, by a God, but then does not interfere whatsoever after that point? It is likely some combination of Deism, Unitarianism and Determinism. I’m sure that Christians would relegate his belief system to the “bottom of the pile” but should he? After all, if there is a God it is likely that there is only one and we would all worship that same God.
So what is this powerful energy generating system?
The Krebs Cycle is now understood much differently than when in Medical School. We learned it as a simple chemical pathway/circle of reactions between compounds that generated energy. It begins with oxaloacetate taking on an acetyl-CoA to form Citrate to begin the cycle. It looked like this:

BUT…now, rather than viewing it as producing ATP, we know it actually produces the energy precursors, which then enter the ETC (electron transport chain) heading out of the inside matrix into and through the intracellular membrane the created protons (H+) and electrons. Then a separate molecule performs “oxidative phosphorylation” as it heads in the opposite direction back into the matrix using the magical enzyme, ATP Synthase, to fix Pi onto ADP to form our ATP:
So, before these complexes can be used, we need the Krebs Cycle no produce high energy molecules.
Glycolysis occurs in the cytosol making pyruvate from glucose (a good Khan Academy video is here). Pyruvate is oxidized into acetyl CoA as it moves into the mitochondrial matrix (deep to the membranes) where it begins in the Krebs cycle, the product of which are substrates NADH and FADH2. Recall that the Krebs cycle (tricarboxylic acid cycle) is a circle that begins with acetyl-CoA combining with oxaloacetate and converted to citrate. There are 8 reactions before getting back to oxaloacetate with enzymes used for each step. All except succinate dehydrogenase (integral to the membrane, the 6th step converting succinate to fumarate) are in the mitochondrial matrix. Krebs itself does not require O2. O2 is the receptor for the electrons coming in through the OxPhos reactions next. It only generates 2 ATPs as opposed to the OxPhos reactions next that generate 36 per molecule of glucose.
Next comes the oxidation and removal of electrons from these precursors.

There are 2 types of protein complexes embedded in the inner-membrane called the ETC (I-IV) and ATP synthase (V).
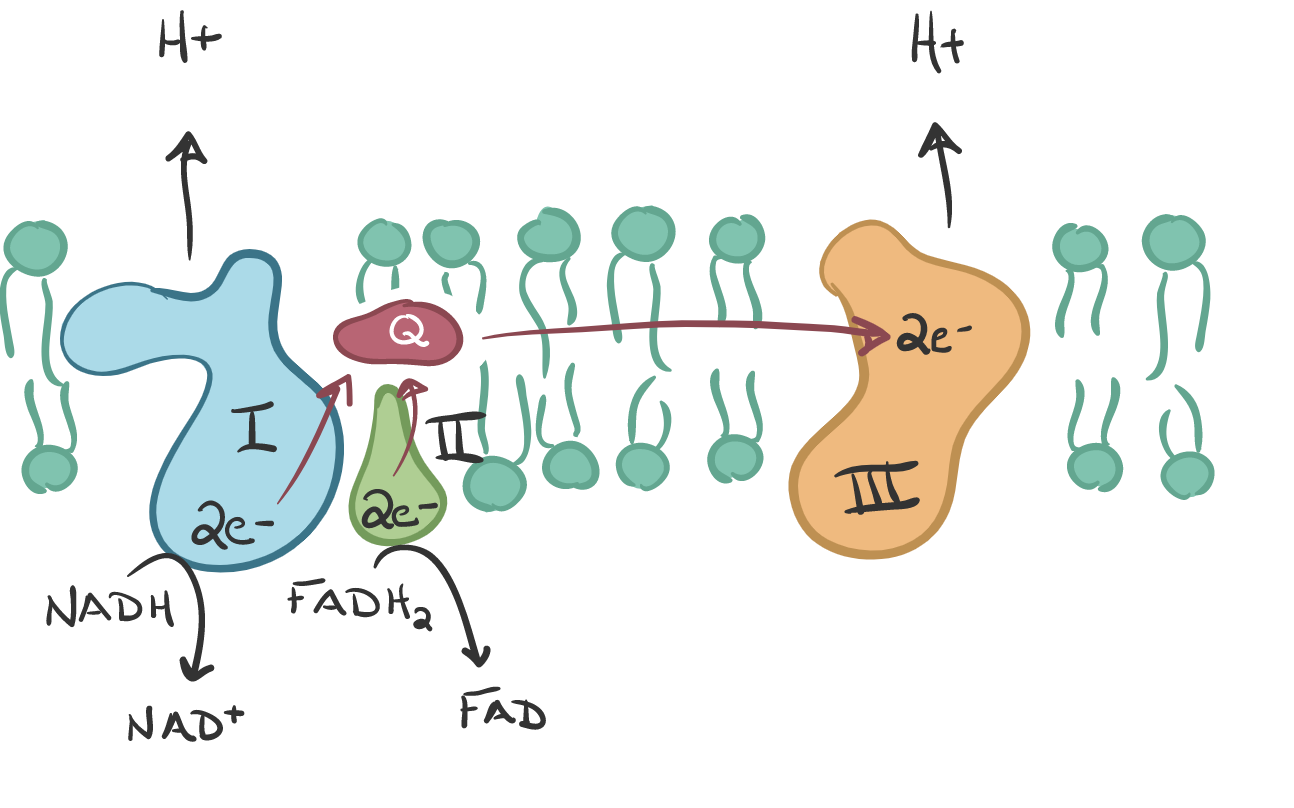
These precursors then enter the Electron Transport Chain, which are 4 complexes, I - IV. The Krebs cycle produces 3 NADH and 1 FADH2. Complex I grabs the NADH high energy molecule and Complex II the FADH2 which causes a loss of electrons from each (oxidation is the loss of electrons). This process forces (pumps) the proton (H+) from the matrix to the intramembrane space. The electrons in Complex I are given to another ‘membrane-bound’ carrier called ubiquinone (Q). It is ubiquitous thus the name.
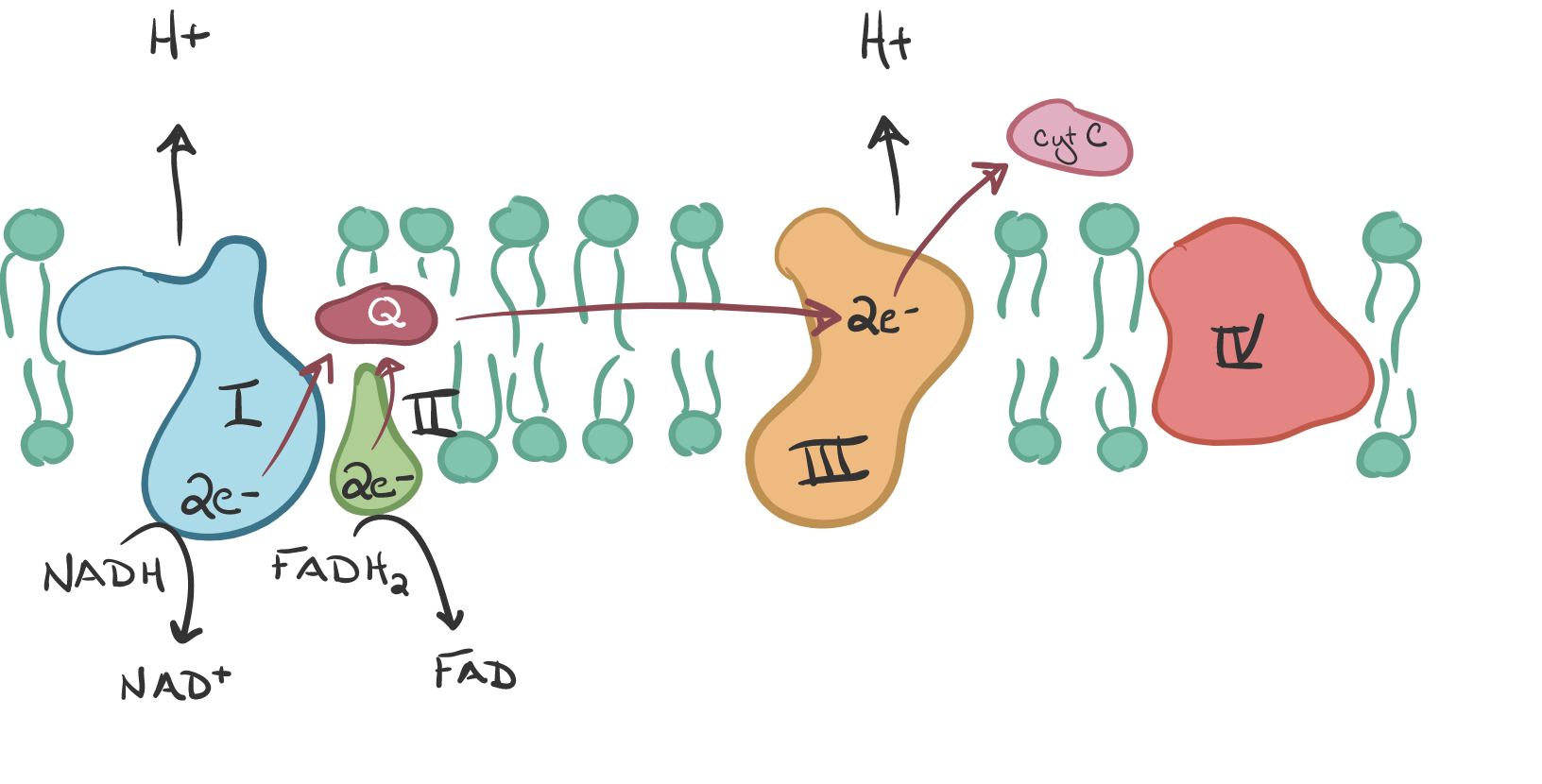
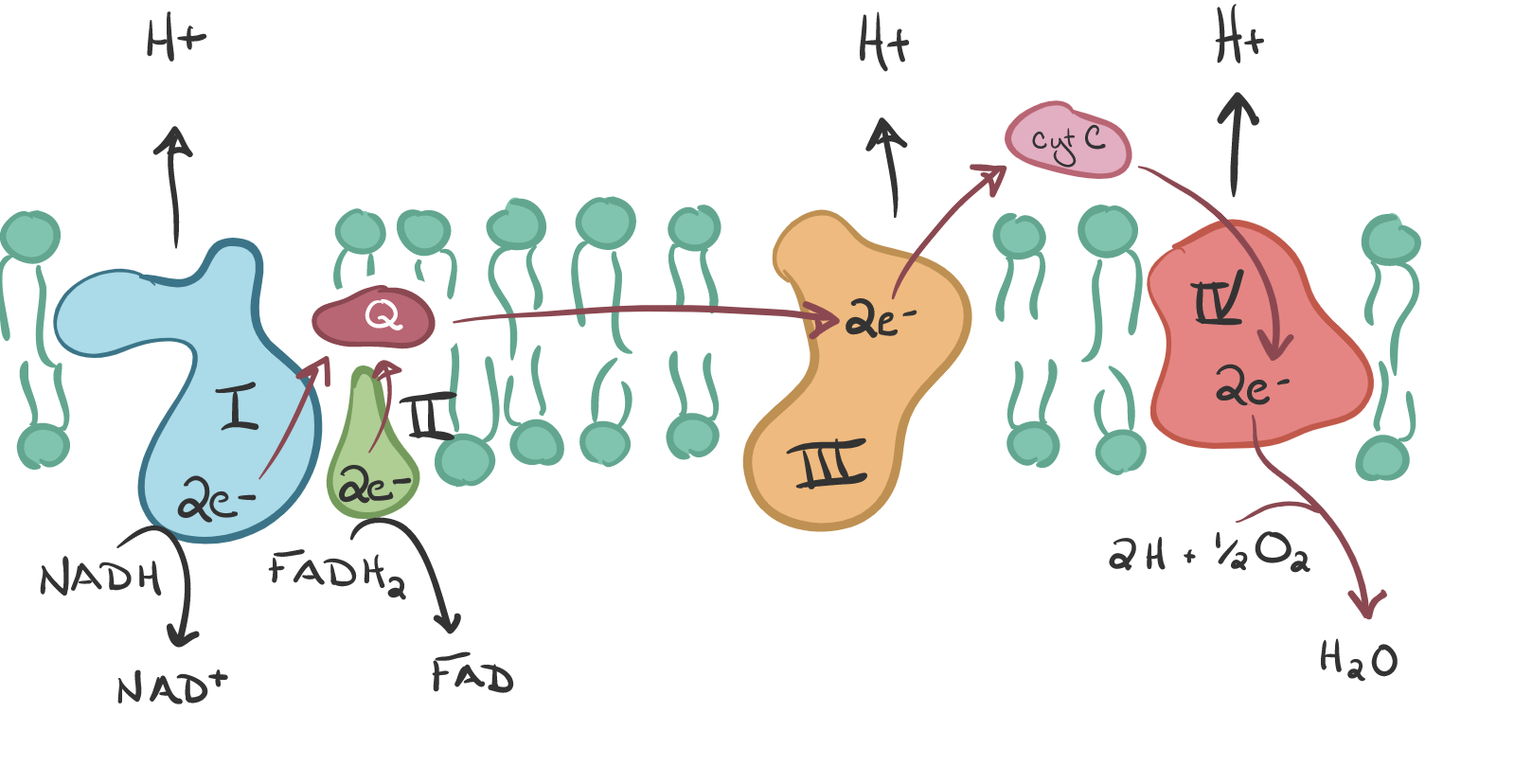
The electrons tat arrived at protein complex 3 are picked up by cytochrome C, the last electron carrier, causing protons to be pumped into the intramembrane space. This transferring of electrons drives the pumping of protons “against gradient” since H+ ions accumulate in the intramembrane space. Thus, energy is stored temporarily. Note that FADH2 behaves differently, transferring its electrons directly to Q with the H2 so no H+ ions are pumped. Also, it uses the succinyl dehydrogenase which is the only enzyme embedded in the membrane.
Cytochrome C carries the electrons to the final protein complex, Complex IV. These electrons are then drawn to O2, the final electron acceptor. O2 must be present for OxPhos to occur. Water is formed as O2 receives the electrons from IV and combines with protons on the inside of the cell.
In summary we are now +3 NADH, +1 FADH2, +3 H+, -2 H+, -1/2 O2 AND +1 H2O.
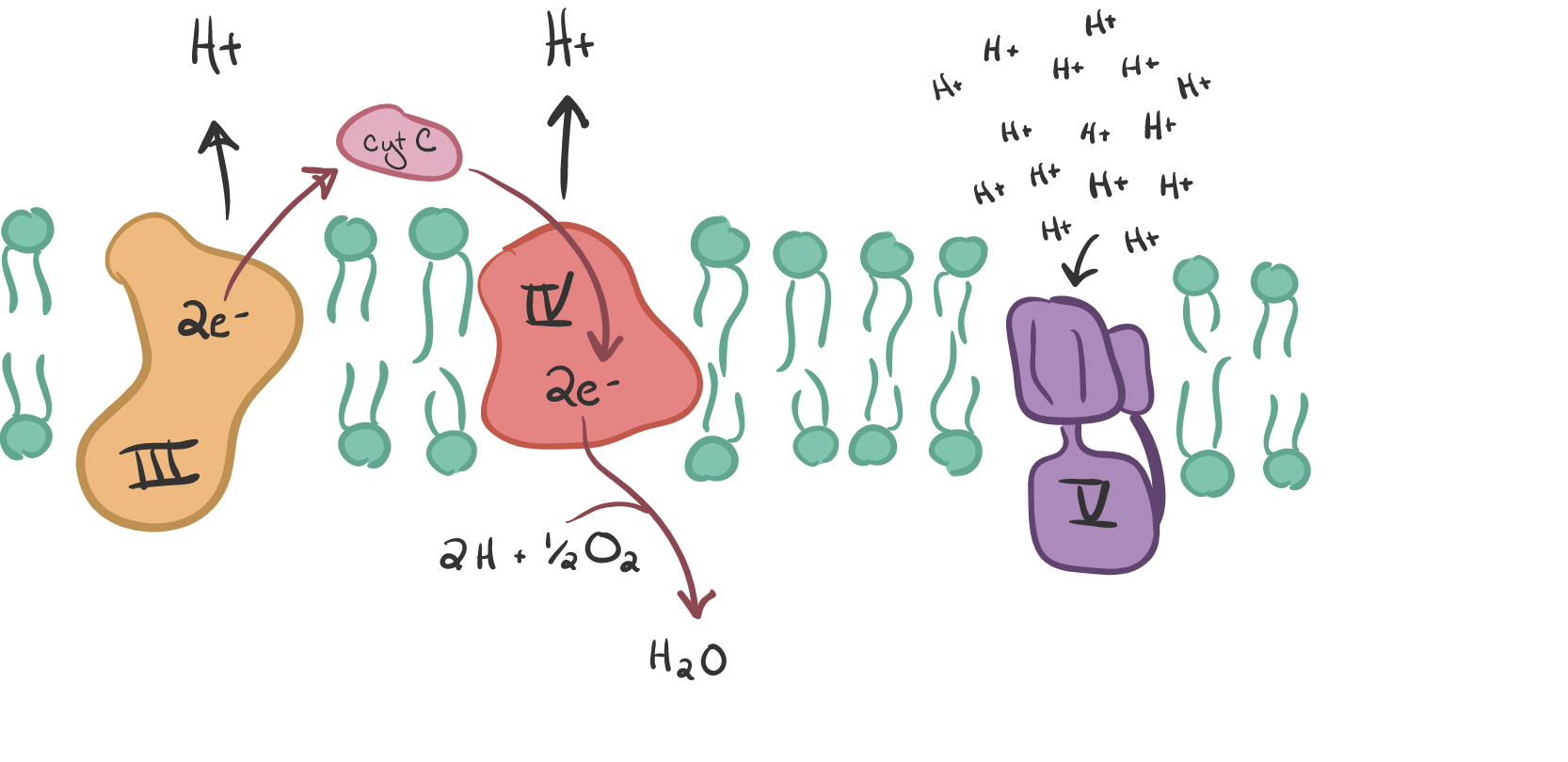
When the H+ concentration is high in the intramembrane space, the ATP synthase comes into action. It is a complex of 2 functional domains, F₁ and F⚬. F₁ is the “rotor” with 5 subunits, 3 each of the alpha and beta (the stator) and one of the gamma, delta and epsilon (the stalk/rotor). The Fo has subunits c, a, b, d, F6, OSCP plus accessory subunits ee, f, g, and A6L. I only mention these units to emphasize the complexity of this system. As the protons pass through Fo, the energy is transferred to F1 where ADP is phosphorylated to ATP. The H+ concentration creates a proton-motive force of a) pH differential and a b) electrical membrane potential.
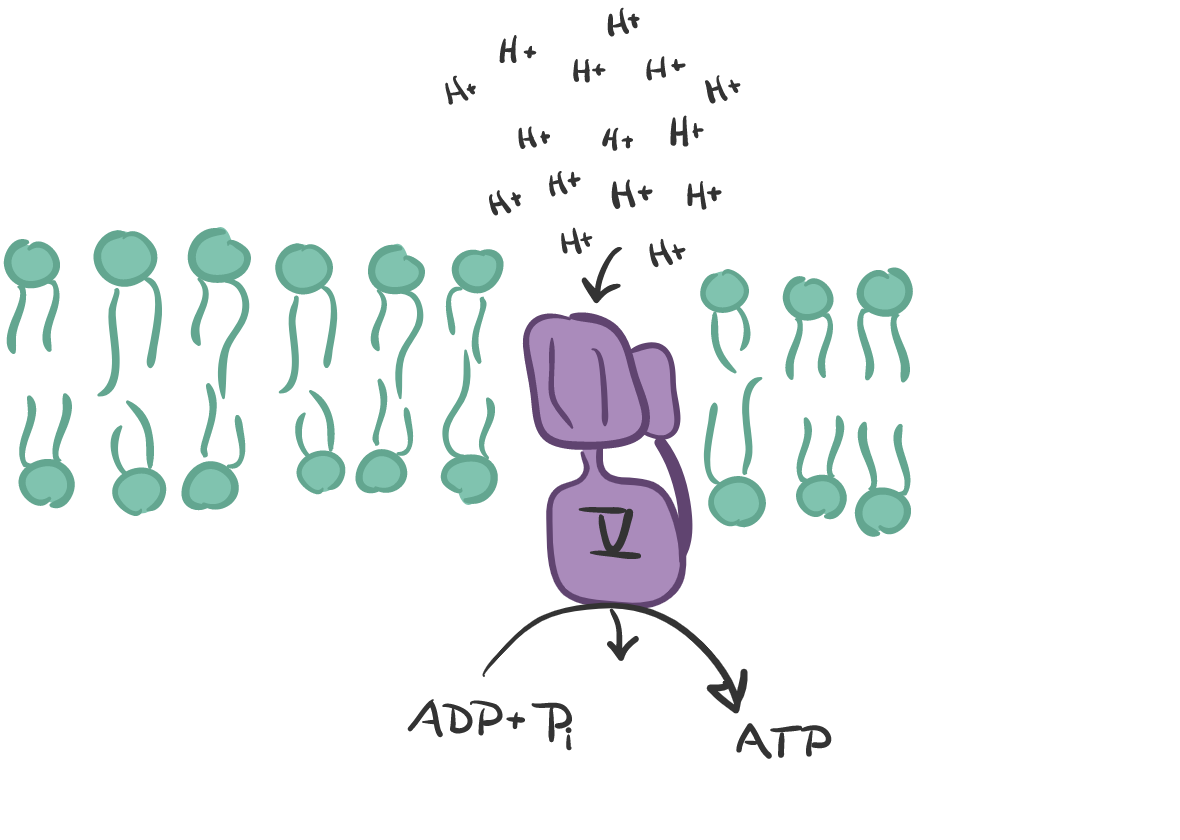
When there is high concentration of protons on the outside of the mitochondrial membrane, protons are pushed through the ATP synthase. This causes ATP synthase to spin, and bind ADP and inorganic phosphorus (Pi) producing ATP…FINALLY! The process of passing H+ through the Complex V is called chemiosmosis. But when it happens in a spinning molecule, it is called substrate phosphorylation. So imagine that this magnificent, elegant and complex machinery was “invented” as life began! It did not “evolve” from a slim-pond struck by lightning or from deep sea thermal vents. There has been no success in demonstrating any way to arrive at this system naturally.
 That’s a pretty heavy introduction at the top and the biochem following is plenty heavy too, at least for me. ; - ) While technically addressed to atheists, I wonder if some others might want to weigh in –
That’s a pretty heavy introduction at the top and the biochem following is plenty heavy too, at least for me. ; - ) While technically addressed to atheists, I wonder if some others might want to weigh in – 