I guess you were never a fan of poetry? I’m being a little figurative with my wording because I haven’t found any writer on this topic who has the benefit of the most technically correct phrases. But let me try to give you a sense of what I mean.
In an earlier thread, you listed links to articles:
Speciation genes (chapter from Encyclopedia of Evolutionary Biology
You introduced these links with the following: “Here are some basic reviews of speciation genes, most of which are genes that somehow influence reproductive isolation.”
So. . . we’ve established that there are genes that “somehow influence reproductive isolation”. Ben, no doubt, would expect many of them to be Alleles. I would too. So, what if you find that these particular genes are especially concentrated in the sub-population with a certain blood type, or with a certain coloration.
Once we have identified which groups have the most potent configurations of “genes for reproductive isolation” . . . then it would be possible to test a hypothesis by introducing known quantities of adults having the Opposite configurations, right?
And the more adults with this “anti-isolation” genetic configuration, the more diluted becomes the population regarding the genetic predisposition that keeps them from successfully mating with animals at the other end of the “ring”.
I’m perfectly open to better terminology … but at the moment, I have to start somewhere. You didn’t like the phrase “Ring Speciation” - - I thought you were being a little “fussy” … it’s just 2 words. But okay … so I’m avoiding using that phase.
But your complaint against “trend” seems a bit more fussy … in that the “trend” in a Ring Species is for the two ends of the “ring” to continue to become more and more different from each other, right? Isn’t that a fundamental truth? As soon as you have two species, the genetic differences between species inevitably grows?
But the Amazing Thing about ring species is that there is still an external genetic vector that can influence the two terminal populations: the adjacent populations in the “ring” ! Unlike all the ordinary species of the world… these adjacent vectors provide a source for “genetic injections” which theoretically allow accelerating or slowing down the ongoing process of speciation!
I hope my comments, as fuzzy as they may seem, are still grammatically correct enough for you to follow my thoughts on the truly unusual nature of Ring Species genetics!
Anyone who understands basic genetics would expect genes of every kind in every population on every planet to have alleles.
That’s just called gene flow. It’s not amazing, it’s a basic pop gen concept. It’s in every intro biology textbook in the chapter on evolution. It happens in all populations all the time.
There is nothing unusual at all about ring species genetics. I can’t tell why you think there is. It seems you have latched onto ring species as a unique or extraordinary phenomenon. The correct way to think about a ring species is that it is a rare and remarkable outcome of basic and very well known population biology. They are rare, not because there is any weird or unique genetic mechanism operating, at least none that we know of or need to imagine, but because the ecological/behavioral combination of conditions is rare.
A very recent paper has explored ring species formation, through modeling. I have only seen the abstract, but you will see that gene flow and habitat structure, along with mobility, are the presumed factors involved.
I tried to explain above what is unusual or different about ring species. It’s not genetics. You can take it from here.
3 Likes
gbrooks9
(George Brooks, TE (E.volutionary T.heist OR P.rovidentialist))
43
I plan to comment on most of your post a little later. But for now, I want to say how brilliant even the Extract of the article you linked for the readers!
Barriers to gene flow and ring species formation.
by de Brito Martins A1,2,3, de Aguiar MA1.
Extract from the Abstract
“They [Ring Species] provide a rare opportunity to understand the role of spatial structuring in speciation. Here, we simulate the evolution of ring species assuming that individuals become sexually isolated if the genetic distance between them is above a certain threshold.”
Great stuff . . . measuring or modeling that threshold is some great science!
“. . . if populations are confined to narrow ranges by geographic barriers, ring species formation increases when local mating is less spatially restricted. Ring species are most likely to form if a population expands while confined to a quasi-unidimensional range but preserving high mobility in the direction of the range expansion.”
Prof. Stephen, I understand your desire to make absolutely clear that I understand that formation of a Ring Species is not some magical genetic transformation. I am absolutely clear on that.
But like anything else, changes in the environment can only shape a gene pool to the extent that the genetics of the population allow for it to happen. One candidate population for Ring Formation might be “loaded” with the genes that appear more involved in speciation than other genes.
While another candidate population might have a paucity of those genes. I am interested in how to apply math to the difference between “loaded” vs. a “paucity” of such genes.
It has been my working hypothesis that the man-made intrusion into Dog gene pools has had the affect (intended or not) of reducing the presence of genes that promote speciation. But now that I have some pretty good articles as benchmarks on speciation, I wont spend that much time on Dogs any further. ( I’m sure some readers will be delighted with the news!)
The Abstract sentence I quote above (**“They [Ring Species] provide a rare opportunity to understand the role of spatial structuring in speciation.”) focuses on the environmental triggers, while I will be focusing my readings on the genetic propensity to respond to such environmental triggers! Obviously, if it was universally accepted that there are no genes with special correlation to speciation, then there wouldn’t be anything to study.
But you yourself have offered the evidence that there are genes that appear to support or promote speciation (all other things being equal).
Formation of Ring Species is a very exciting topic!
Stephen, more later in response to your post.
gbrooks9
(George Brooks, TE (E.volutionary T.heist OR P.rovidentialist))
44
Wow!
It would seem that a little knowledge can either be a dangerous thing … or it could lead to more knowledge!!!
@sfmatheson and @Lynn_Munter, I’m sure you will find this discovery either wonderful or bewildering. For sure some some readers will be pretty danged surprised to find the phrase “Incipient Ring Speciation” in a scientific journal article! In fact, in Darren Irwin’s article (“A Novel Approach for Finding Ring Species…”), we find the phrase “Incipient Ring Speciation” twice! (once in the body and once again in a footnote).
But just so nobody starts to yawn … we also find just the two words (“Ring Speciation”) three times as well !!! Here’s the context for two of those uses!:
"Given the very large number of identified candidate barriers, it is almost
inevitable that at least one will be associated with any interesting species
complex that we might point to as a candidate for ring speciation, <<<< [See what he did here!]
and this means that the predictive value of the model will depend on further
refinement. Despite these issues, it is likely that the present model represents
an important first step in this geography-oriented approach to the analysis of barriers involved in both ring speciation <<<<<<<<<<<<<<<<<<[Can you believe he did it again ?!?]
and speciation more generally. "
National Center for Biotechnology Information, U.S. National Library of Medicine
8600 Rockville Pike, Bethesda MD, 20894 USA
Journal List > BMC Biol > v.10; 2012 > PMC3299606
BMC Biol. 2012; 10: 21.
Published online 2012 Mar 12. doi: 10.1186/1741-7007-10-21
A novel approach for finding ring species: look for barriers rather than rings
Darren E Irwin, corresponding author
Abstract
Ring species, in which two different forms coexist in one region while being connected by a long chain of interbreeding populations encircling a geographic barrier, provide clear demonstrations of the evolution of one species into two. Known ring species are rare, but now Monahan et al. propose an intriguing new approach to discovering them: focus first on geography to find potential barriers.
See research article
Commentary
Speciation, the process by which a single species evolves into two or more, is difficult to observe directly because of the long span of time it usually takes to occur. Nonetheless, biologists have been able to infer much about speciation by examining geographic variation within and between species. A striking pattern that emerged about a century ago is known as Jordan’s law [1]: given any species, the most closely related species is found ‘in a neighboring district separated from the first by a barrier of some sort or at least by a belt of country, the breadth of which gives the effect of a barrier.’ The role of such barriers in speciation is perhaps best illustrated by the rare phenomenon known as ‘circular overlaps’ [2] or ‘ring species’ [3], when two coexisting but reproductively isolated forms are connected by a long chain of populations encircling a geographic barrier, and traits change gradually from those of one form to the other around the ring [4] (Figure (Figure1).1). The great evolutionary biologist Ernst Mayr called such situations the ‘perfect demonstration of speciation’ [2] since they allow one to use geographic variation to infer how evolutionary change in time led to the differences between species.
Map of the geographic distribution of an idealized ring species. Two forms (red and blue; species A and B) have come into contact (perhaps with some overlap) but do not interbreed directly. They are connected by a long chain of populations encircling …
Until now, our knowledge of the diversity of ring species has arisen primarily from the field of taxonomy, with experts on the taxonomy of particular groups occasionally noticing a pattern of gradual variation between quite divergent forms. This somewhat haphazard approach has led to a variety of ring species being proposed [2,4], only some of which have held up to further scrutiny [4,5]. Only two well-studied cases are generally accepted as solid examples of ring species: these are the Ensatina eschscholtzii salamander complex in California [6] and the Phylloscopus trochiloides greenish warbler complex in Asia [7]. One challenge in relying on taxonomists to discover ring species is that the naming rules of taxonomy generally conceal their existence: taxonomists have to decide whether a group of specimens is two species or one species; the taxonomic naming system does not lend itself toward describing gradients between two species [4].
The study by Monahan et al. [8] proposes a novel approach to the discovery of ring species, focusing on geography rather than taxonomy as the starting point. They ask an intriguing question: where in the world are there barriers that might promote ring speciation? A topographic model, based on slope of the landscape, is used to identify potential geographic barriers worldwide. In the model, barriers are regions that have either more or less slope than the regions around them. The characteristics of the potential barriers, such as size and shape, are then compared with those of known barriers in two ring species (E. eschscholtzii salamanders and P. trochiloides greenish warblers) and two groups that have been proposed as ring species and share many of their characteristics (Acacia karoo trees and Larus gulls). Known barriers are similar to only a small proportion of all potential barriers, suggesting that ring species barriers have common characteristics. The authors also show maps of a small subset of the potential barriers that are similar to the real ring species barriers, suggesting that these may be good locations to look for ring species.
Though the current model is based solely on slope, other geographic and environmental variables could eventually be incorporated to enhance the effectiveness of the model in identifying some barriers in species distributions. In particular, it may be advantageous to introduce elevation as a geographic variable in the model. The current use of slope results in two sorts of ‘barriers’ being identified: 1) areas of high slope, such as mountain ranges, escarpments, or ocean trenches, surrounded by areas of low slope such as plains, plateaus, or ocean basins; and 2) areas of low slope surrounded by those of high slope. As a result, some of the barriers identified by this model are peculiar: for example, in the first case, an area of flat land bordered on one side by a steep climb toward higher elevations and on the other side by a steep drop toward lower elevations; in the second case, a steep escarpment between a high plateau and a low plain. In both of these, it seems unlikely that a species could live in all areas encircling the ‘barrier’ without also inhabiting the ‘barrier’ itself. Rather, it seems that the optimal topographic model would use some combination of both slope and elevation to identify barriers. Elevation is also likely to work better than slope in describing the Arctic Ocean barrier in the case of the Larus gull ring; the slope-based model results in three separate barriers corresponding to deep ocean basins, which the authors then joined as a composite barrier (see [8], their Figure 2D). It seems that slope on the deep ocean floor is of little relevance to describing the distribution of a bird species, whereas elevation (for example, above or below sea level) is of substantial importance.
Environmental variables such as climate or vegetation could also be incorporated into the model. For instance, with respect to the central Asian barrier that the greenish warbler encircles, Monahan et al. find that their model did not identify a single barrier - rather, they construct a composite barrier out of two separate barriers identified by the model. They remark that, in cases such as this, ‘it is difficult to imagine any univariate or multivariate environmental approximation of a single barrier (for example, Central Asia, which is comprised of the Takla Maka-Gobi deserts and the Tibetan Plateau - large geographic regions that differ dramatically in terms of climate and vegetation).’ However, a good explanatory variable has been identified in this case: greenish warblers inhabit forests [7], and maps of forests in Asia (for example, [9]) show a large gap that includes the Tibetan Plateau as well as the Taklamakan and Gobi deserts. Other examples of large potential barriers that show up clearly when considering a basic environmental variable (wet versus dry) are Antarctica, Australia, and Greenland (for marine and/or terrestrial coastal organisms), which were missed by the current topographic model. It is clear that the addition of other topographic and environmental variables could greatly enhance the precision of the model, and Monahan et al. [8] emphasize that their general approach can be modified to work with any kind of continuously distributed environmental variable, making it of wide applicability to many different types of investigations into barriers to dispersal that may contribute to speciation.
Finally, the very large number of potential barriers identified by the topographic model (952,147, about 10,000 of which are ‘topographically similar’ to those associated with known ring taxa [8]) raises another issue. Given the very large number of identified candidate barriers, it is almost inevitable that at least one will be associated with any interesting species complex that we might point to as a candidate for ring speciation, and this means that the predictive value of the model will depend on further refinement. Despite these issues, it is likely that the present model represents an important first step in this geography-oriented approach to the analysis of barriers involved in both ring speciation and speciation more generally. The approach proposed by Monahan et al. [8] will likely be adapted to incorporate multiple variables (in addition to slopes), and this will allow more refined identifications of a smaller number of potential barriers, resulting in more useful predictions. The discovery and inclusion of more ring species (for example, the willow warblers Phylloscopus trochilus, which display a form of incipient ring speciation around the Baltic Sea [5,10]) will likewise allow further refinement of the model, perhaps eventually allowing an analysis of what types of barriers are associated with ring species from different taxonomic groups. By applying an explicit geographic framework to the analysis of ring species, Monahan et al. have pioneered an interesting new approach to the study of the relationship between geography and speciation. In the years ahead, it will be exciting to see whether additional ring species are identified using this geography-oriented approach.
References:
1.Jordan DS. The law of geminate species. Am Nat. 1908;42:73–80. doi: 10.1086/278905. [Cross Ref]
2.Mayr E. Systematics and the Origin of Species. New York: Dover Publications; 1942.
3.Cain AJ. Animal Species and their Evolution. London: Hutchinson House; 1954.
4.Irwin DE, Irwin JH, Price TD. Ring species as bridges between microevolution and speciation.
Genetica. 2001;112-113:223–243. [PubMed]
5.Irwin DE. Incipient ring speciation revealed by a migratory divide. Mol Ecol. 2009;18:2923–2925. doi: 10.1111/j.1365-294X.2009.04211.x. [PubMed] [Cross Ref]
6.Wake DB. Incipient species formation in salamanders of the Ensatina complex. Proc Natl Acad Sci USA. 1997;94:7761–7767. doi: 10.1073/pnas.94.15.7761. [PMC free article] [PubMed] [Cross Ref]
7.Irwin DE, Bensch S, Price TD. Speciation in a ring. Nature. 2001;409:333–337. doi: 10.1038/35053059. [PubMed] [Cross Ref]
8.Monahan WB, Pereira RJ, Wake DB. Ring distributions leading to species formation: a global topographic analysis of geographic barriers associated with ring species. BMC Biol. 2012;10:20. [PMC free article] [PubMed]
10.Bensch S, Grahn M, Müller N, Gay L, Åkesson S. Genetic, morphological, and feather isotope variation of migratory willow warblers show gradual divergence in a ring. Mol Ecol. 2009;18:3087–3096. doi: 10.1111/j.1365-294X.2009.04210.x. [PubMed] [Cross Ref]
gbrooks9
(George Brooks, TE (E.volutionary T.heist OR P.rovidentialist))
45
Then, of course, we have the article that uses the phrase “Ring Speciation” in its Title !!!
Incipient ring speciation revealed by a migratory divide.
by Irwin DE
Mol Ecol. 2009 Jul;18(14):2923-5. doi: 10.1111/j.1365-294X.2009.04211.x. Epub 2009 May 7.
Abstract
Ever since Ernst Mayr (1942) called ring species the ‘perfect demonstration of speciation’, they have attracted much interest from researchers examining how two species evolve from one. In a ring species, two sympatric and reproductively isolated forms are connected by a long chain of intermediate populations that encircle a geographic barrier. Ring species have the potential to demonstrate that speciation can occur without complete geographic isolation, in contrast to the classic model of allopatric speciation. They also allow researchers to examine the causes of reproductive isolation in the contact zone and to use spatial variation to infer the steps by which speciation occurs.
According to the classical definition, a ring species must have (i) gradual variation through a chain of populations connecting two divergent and sympatric forms, and (ii) complete or nearly complete reproductive isolation between the terminal forms. But evolutionary biologists now recognize that the process of speciation might often occur with some periods of geographic contact and hybridization between diverging forms; during these phases, even partial reproductive isolation can limit gene flow and permit further divergence to occur.
In this issue Bensch et al. (2009) make an exciting and important contribution by extending the ring species concept to a case in which the divergence is much younger and not yet advanced to full reproductive isolation. Their study of geographic variation in willow warblers (Phylloscopus trochilus; Fig. 1) provides a beautiful example of gradual variation through a ring of populations connecting two forms that are partially reproductively isolated where they meet, possibly due to divergent migratory behaviours of the terminal forms.
gbrooks9
(George Brooks, TE (E.volutionary T.heist OR P.rovidentialist))
46
In one of the articles I have posted already, there was this fascinating quote:
“Ever since Ernst Mayr (1942) called ring species the ‘perfect demonstration of speciation’, they have attracted much interest from researchers examining how two species evolve from one. . . . Ring species . . . demonstrate that speciation can occur without complete geographic isolation, in contrast to the classic model of allopatric speciation.”
Who was this German-born scientist, Ernst Mayr? I was pretty surprised that he is the person who actually taught new generations of geneticists that the identification of a species should not be based purely on differences in behavior or appearance. He argued, as early as his 1942 book, “Systematics and the Origin of Species”, that species should be defined by their reproductive incompatibilities!
The irony of this assertion was that it brought (intentionally or unintentionally) the Biblical notion of a “kind bringing forth it’s own kind” into the scientific discussion !
“When populations within a species become isolated by geography, feeding strategy, mate choice, or other means, they may start to differ from other populations through genetic drift and natural selection, and over time may evolve into new species. The most significant and rapid genetic reorganization occurs in extremely small populations that have been isolated (as on islands).”
This posting is a salute to my new personal hero … he lived to the age of 100, dying only 12 years ago, and writing books from 1942 to just a year before his death!
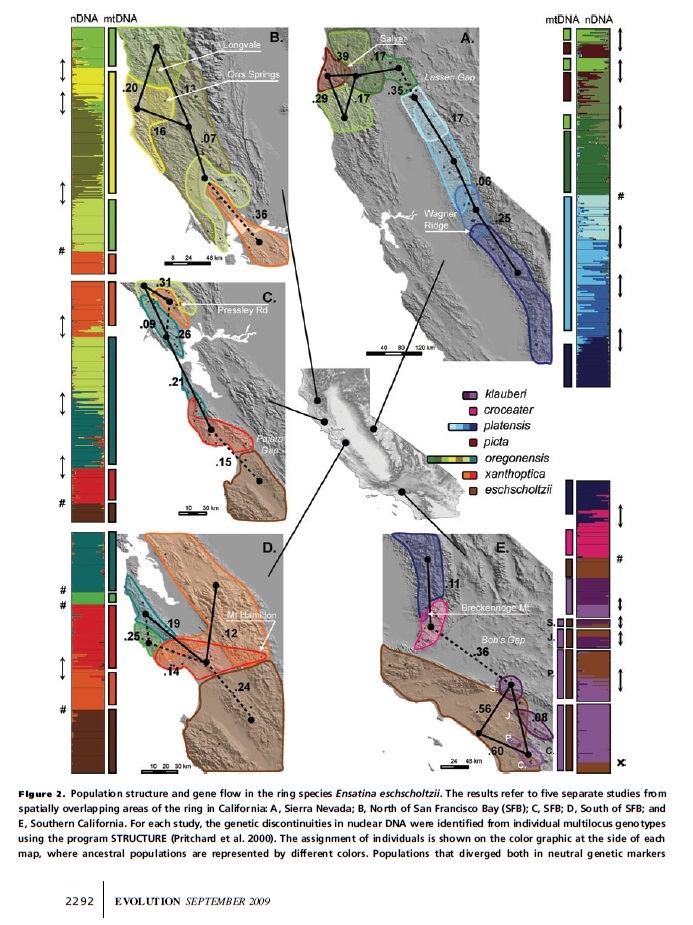
GENETIC LEAKAGE AFTER ADAPTIVE AND NONADAPTIVE DIVERGENCE IN THE ENSATINA ESCHSCHOLTZII RING SPECIES by Ricardo J. Pereira1,2,3,4 and David B. Wake1
1Museum of Vertebrate Zoology and Department of Integrative Biology, 3101 Valley Life Sciences Building, University of California, Berkeley, California 94720
2E-mail: ricardo@berkeley.edu