I would like to ask the help of this group to evaluate whether the origin of the vertebrates is a case of detectable divine intervention. Feel free to add additional scientific information in order to inform this discussion.
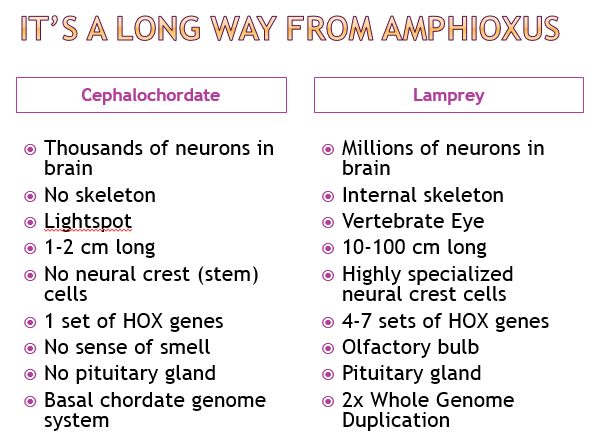
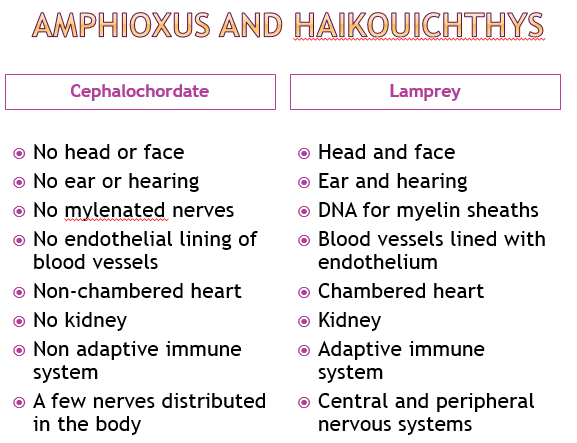
The first animal life in the fossil record is 580 Ma, the Avalon Explosion. A vertebrate lamprey fish appears in the fossil record (chordate) in the Chengjiang Lagerstatten (517 Ma). DNA molecular analysis indicates that a vertebrate fish could not evolve within 60 million years. A cephalochordate (Cathaymyrus) appears in the same Chengjiang formations. Vertebrates are strongly linked to cephalochordates such as amphioxus by physiology and DNA. The lamprey larvae is similar to adult cephalochordates. Most of the vertebrate novelties are formed by neural crest cells, which are lacking in cephalochordates.
My hypothesis is that God modified a cephalochordate by with neural crest cells in order to form the ancestor of all vertebrates.
I made a table of some of the differences between cephalochordates (amphioxus) and the lamprey fish that was found at Chengjiang (Haikouichthys).
An image of Cathaymyrus from Wikipedia (Apokryltaros):
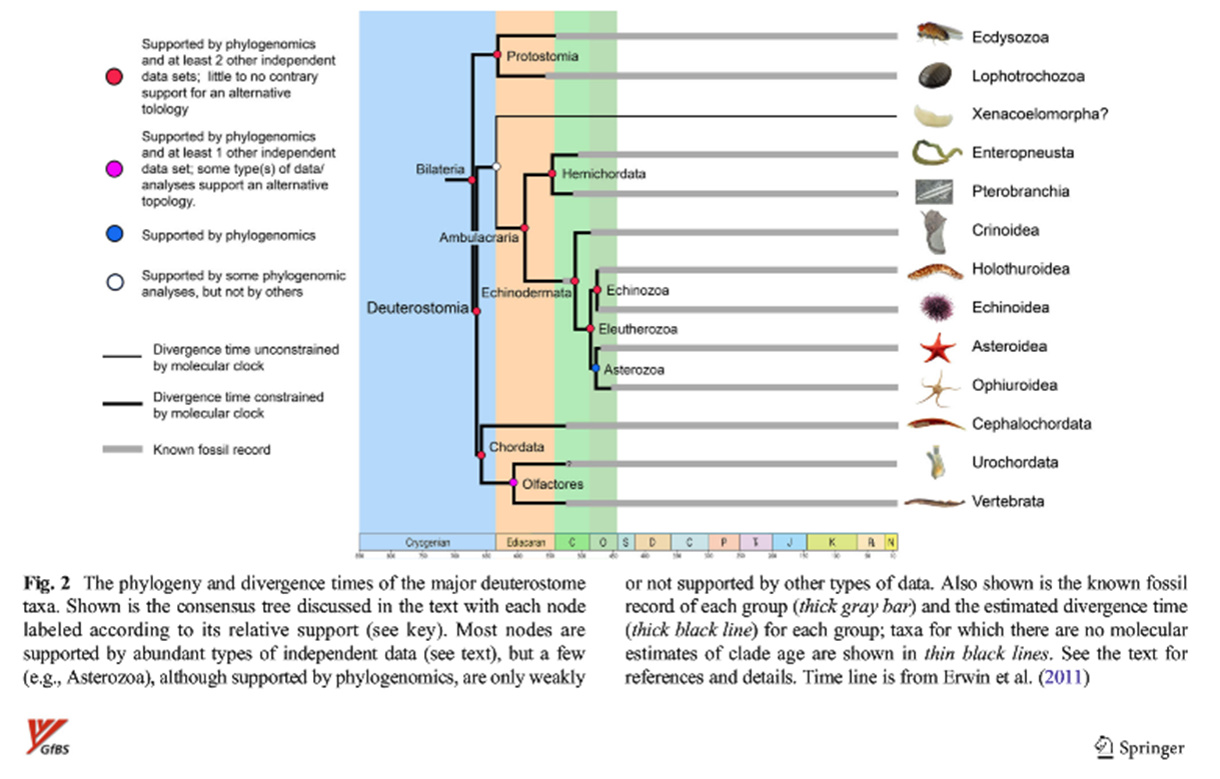
{kind=link}