Do you plan to repeat this statement another 100 times in hope that your audience will have passed away and it will seem new and credible to a brand new bunch?
Natural Selection can be QUANTIFIED, in reference to any reasonably defined time interval, by quantifying the number of offspring surviving to “first offspring” produced by a given type of individual or a given sub-group of a population.
It is no more tautological than any linguistic term might be tautological - - as demonstrated by using a dictionary to define a term, then getting the dictionary definition of the terms used in the first definition and so on.
How far have you studied any particular science you prefer to mention? Ph.D?, Masters? BS or BA? Or even fewer years? Your mastery of linguistic foibles seems to be far and away more advanced than your mastery of how natural objects and natural processes actually work in the world.
T_aquaticus
(The Friendly Neighborhood Atheist)
182
I think it is more of an issue of transparency. Gene sequences for many species can be found online in giant databases. You could even sequence those genomes yourself with a small lab setup and a modest research grant. A 1,000 bp sequencing run is like $5 at most commercial operations, or even cheaper with bulk submission. The idea that scientists are faking the sequences they submit to databases and in publications is not worth considering.
That leaves us with the fact of the data probably being real, and in that case evolution along with common ancestry is the unavoidable conclusion. As @glipsnort will attest to, biology is a messy process so there will never be perfect phylogenies or perfect correlations, but that is expected. The ratio of signal to noise is a scientific principle as old as science itself, and in the case of biology the phylogenetic signal stands heads and shoulders above the noise created by biological messiness (e.g. homoplasies, incomplete lineage sorting, lateral genetic transfer, deep nodes lacking genetic data).
If you want me to quote people with PHDs.. I can do that.. will that suffice?
Let me correct you on something anyway-
Natural selection cannot be quantified. Fitness can be.. How is fitness measured? In terms of Surviving offspring.
What is natural selection ? The fact that the more fit survive.
Though this fact is not always true.. when the less “fit” survive.. its genetic drift…
what are you left with? meaningless drivel that can be interpreted any way thats convenient and never really be falsified..
And thats why tautologies are problematic in science.
A biologists discussion of the problem is given below - Reconsidering the logical structure of the theory of natural selection - PMC
Let me quote critical segments
Blockquote
Tautologies and circular statements are always true and are thus not useful as theories because they can’t be falsified. A tautology is a statement that is true by necessity.4 Tautologies often take the form “a equals b,” but b reduces to a, so really “a equals a.” Similarly, circular statements include components that rely on each other mutually for their definition, taking the general form “a is true because b is true and b is true because a is true.” Tautologies and circular statements cannot be falsified, because they are always true, so they are not scientific under a Popperian view of science.5,6
Blockquote
Darwin first described natural selection in The Origin of Species in a way that is tautological. The full title of his seminal book is On the Origin of Species by Means of Natural Selection, or the Preservation of Favored Races in the Struggle for Life. The sub-title is itself a tautology because “natural selection,” “preservation of favored races” and “struggle for life” all reduce to the same concept, if we accept that “struggle” includes reproduction as well as survival. A equals a equals a. Tautologies can sometimes be useful in terms of describing the same concept with different words that give more information about key ideas (I just did it by using the terms “concept” and “ideas,” which are exactly the same thing in this context). However, in describing the prevailing theory of evolution, we must avoid tautology at all costs because tautology renders the theory empty.
Blockquote
For a more modern description of natural selection, Ayala (2008)7 writes: “Natural selection – i.e. differential multiplication – can accomplish adaptation because a favorable mutation that has occurred in one individual may thus spread to the whole species in a few generations…” Parsing this sentence, we see that “Natural selection,” “differential multiplication,” “favorable,” “accomplish adaptation,” “may thus spread,” are all ways of saying the same thing: more offspring are produced in some situations. Even more simply, all these phrases say, essentially, is that there is some biological change occurring, with no explanation as to how. A equals a equals a equals a equals a. The “how” is what natural selection theory is supposed to provide, but providing mutually inter-defined terms does not do so.
Blockquote
To most encountering this critique for the first time, there is a shake of the head and a refusal to acknowledge that such a serious problem could have persisted in evolutionary theory for this long. It seems that the root of this (wrong) conclusion is the deceivingly common-sense feeling that adaptations and natural selection are not the same concept. They are indeed different in some ways, but they reduce to the same concept. We know intuitively that an opposable thumb, for example, is an adaptation to our environment because it’s so useful in our normal (human) lives. And we know that being able to run faster than potential predators is an obvious adaptation for a deer, through observation. So of course these types of adaptations will spread through the population in a process that we label “natural selection;" that is, nature, as the sum of various forces in the environment of the organism or population at issue, favors (selects) these adaptations.
Blockquote
Lipton and Thompson (1988)10 attempt a more spirited defense of natural selection against charges of circularity by arguing that natural selection explanations are recursive rather than circular and are a type of filter explanation. For example, to answer the question “why are polar bears white?” a filter explanation says “because there is a filter that makes them white.” The filter is then to be specified through examination of the environment and polar bear populations. Lipton and Thompson disavow the very common practice of defining natural selection or adapation in reference to fitness because they recognize explicitly that such explanations are indeed circular and thus empty. But when we examine their attempt to sidestep circularity or tautology by appealing to recursivity and filter explanations we see that filter explanations, as a category, a meta-theory, reduce to a simple posit that there is some natural cause (the filter) that led to the traits at issue. And positing that there is some natural cause for the trait at issue is not helpful as a theory, other than to make a basic distinction between natural selection and supernatural theories of evolution. This distinction itself is important, to be sure – particularly in the context in which Darwin wrote – but it does not qualify natural selection as a general theory without a great deal more development. Lipton and Thompson themselves discuss how theories can be over-general because they don’t always exclude possible explanations, but their own filter theory explanation of natural selection is itself far too underspecified for the reasons just stated. To be useful as hypotheses, filter explanations must specify the details of the filter posited.
Blockquote
These are not entirely new arguments. Numerous writers have criticized natural selection as a tautology or circular argument, including Butler (1882),11 T. H. Morgan,12 Waddington (1960),13 MacBeth (1971),14 Bethell (1976)15 and more recently Rosenberg and Bouchard (2008).16
The tautology/circularity charge, if true, is fatal to natural selection as a theory of how biological change occurs. This is an exceedingly nuanced issue yet it is surprising that so few biologists and philosophers of biology have tackled the issue head on. I suggest in the next section a possible resolution.
Blockquote
Brandon and Ramsey (2007)17 defend the propensity interpretation of fitness, which was developed specifically to resolve the tautology problem:
The prime motivation [for introducing the propensity interpretation] was to make room for an explanatory theory of natural selection, which is tantamount to solving the so-called “tautology problem.” This problem arises from a casual inspection of the phrase “survival of the fittest” and then asking what defines the fittest. If the answer is those that reproduce the most, then it seems we are explaining a phenomenon, differential reproduction, in terms of itself, which is no explanation at all.”
Blockquote
Summing up: we are saved from tautology by the propensity interpretation(s), and its reliance on “expected fitness,” but with a cost: we must acknowledge that there is no a priori property or value of fitness or expected fitness – as Gould and others have argued. The way in which we define fitness or expected fitness is necessarily arbitrary. This is a substantial but not a fatal problem, so we have made progress. In dealing with arbitrariness, we simply need to be clear how we are defining fitness or expected fitness in each circumstance, allowing for useful comparisons across different studies, as well as conceptual coherence. Moreover, for natural selection to earn its reputation as a powerful theory of how evolution occurs, the theory must be developed far more than it has to date, with a focus on testing and improving evolutionary predictions.
Blockquote
Summing up: we are saved from tautology by the propensity interpretation(s), and its reliance on “expected fitness,” but with a cost: we must acknowledge that there is no a priori property or value of fitness or expected fitness – as Gould and others have argued. The way in which we define fitness or expected fitness is necessarily arbitrary. This is a substantial but not a fatal problem, so we have made progress. In dealing with arbitrariness, we simply need to be clear how we are defining fitness or expected fitness in each circumstance, allowing for useful comparisons across different studies, as well as conceptual coherence. Moreover, for natural selection to earn its reputation as a powerful theory of how evolution occurs, the theory must be developed far more than it has to date, with a focus on testing and improving evolutionary predictions.
In short.. stop telling made up stories using NS as a tautology and do some actual predictable science…
Something yet to happen in the way biologists use NS.
The conclusion is interesting:
Blockquote
Developing natural selection as a theory will require developing a detailed set of filter theories for particular circumstances. These sub-theories will, after testing in numerous experiments, eventually allow development of what may be labeled “principles of evolution.” The good news is that increasingly sophisticated computer models are being developed that should, with good empirical data, allow for the development of the suggested principles more readily than without these powerful new tools. It may be the case, however, that many aspects of evolution in actual populations will remain forever unknowable due to the inherent complexity in such populations. Once we are able to develop models for accurately predicting the course of evolution in actual populations (if not all), natural selection may fairly be described as a general theory of evolution.
While the details of the proposed principles of evolution are beyond the scope of this paper, and certainly beyond my current knowledge, it does seem clear that a new synthesis of evolutionary theory will be far more pluralist than today’s focus on adaptation and natural selection would suggest. I have proposed the outlines of a Generalized Agentic Selection theory as a complement to natural selection in Hunt (2011).20 An in-progress paper fleshes out these ideas. The present critique refers only to natural selection and doesn’t delve into the issues surrounding the origin of variation. The Modern Synthesis posits that variation is random, but it seems that an increasing number of non-random sources of variation, such as Shapiro’s21 natural genetic engineering, or Kirschner and Gerhart’s “facilitated variation,”22 must also be considered if we are to create a more predictive theory of evolution.
Let the referred papers be further reading material…
Edit: Another thing i hate about discussions of these things at popular levels.. People who defend evolution resort to weak arguments and name calling… while actual scientists ae seriously discussion these issues and do accept that the criticism has merit.
I found a more systematic argument in a published scientific papers on this subject from the other side(i.e that evolution happened, but cannot be represented by a tree of life). The basic argument is that that the tree of life is a forced construct and not “real”. It touches most of teh important points and is a structured argument.
The paper is below - http://www.pnas.org/content/104/7/2043
I will quote the paras which will give a gist of the argument (you can read the paper for a detailed understanding)
Criteria for falsifying the tree of life -
Blockquote
The TOL hypothesis could be falsified by substantial failure of any of these propositions. First, and most fundamentally, the pattern of groups subordinate to groups might be illusory or artifactual, “the mere putting together and separating objects more or less alike” (16) in accordance to expectation. In this case there would be no explanandum, no all-embracing pattern or fact existing in nature and independent of our desire to impose order. Second, similarities between species used to erect the TOL (or any natural scheme) might not predominantly reflect common descent. Patterns of resemblance recognized by systematists could after all result from some natural cause other than direct (branching) descent with modification, such as environmental constraint and convergence, parallelism, or reticulation. Third, selection and branching species divergence might not be inevitably connected. Sometimes selection will drive reticulation [as with lateral gene transfer (LGT) of novel adaptations], whereas sometimes divergence will be produced by stochastic processes (drift).
What will be argued for:
Blockquote
We will argue that inclusive hierarchical classifications do not emerge naturally and consistently from the relevant prokaryotic data considered in general (in their entirety). Instead, they have been imposed on them by selective analyses that are based on the assumption that a tree must be the real natural pattern, even if only certain of the data can be trusted to reveal it. Furthermore, we propose that the underlying historical processes affecting prokaryotes are more complex and various than those imagined by Darwin (or by neo-Darwinists), and not of necessity expected to give rise to a natural hierarchy.
Being able to create trees is not the proof for Trees of Life -
Blockquote
The body of data (the explanandum) for which a hypothesis (the explanans) proposes to account cannot at the same time constitute proof for that hypothesis (17, 23), nor can further data of the same kind. We might construct a hierarchical taxonomy of Drosophila based on certain morphological characters and claim that its branching pattern reflects an evolutionary branching process. Adding more taxa would bush out the tree but not strengthen this fundamental claim about process, nor would adding more characters, necessarily, if there were reason to believe that by functional constraint these characters were correlated with the first set. Much of what has happened in post-Darwinian phylogenetics has been an enormous expansion of the explanandum (accepted from the outset by Darwin) by the addition of new taxa or characters (24–32). Moreover, this expansion has for the most part used algorithmic tools that are constrained to produce trees.
Trees are assumed… and not proven -
Blockquote
In a search for independent evidence of a natural hierarchy, Panchen considers homology, paleontology, and biogeography. The first is problematic in that true (taxic) homologies cannot be distinguished from false ones (homoplasies) without some assumption of hierarchy: homologies are more often deduced from trees than trees are from homologies. Thus, explanans melds with explanandum, and neither is tested. The second and third may offer independent evidence that evolution by descent with modification has occurred but are limited in their relevance and applicability to specific groups, areas or times. They do not justify, except by extrapolation, the expectation that there should be groups under groups at all levels, that there should be a universal TOL, dichotomously branching all of the way down to a single root. Alternatives [extensive reticulation or separate origins from a common inchoate ancestral state (2, 14, 34)] can be entertained.
This is more or less what i am saying about convergence / homologs and homplasies.
4, Molecular Phylogenetics in Prokaryotes and implications -
Blockquote
Because there is substantial disagreement among prokaryotic molecular data sets and little strongly supported congruent signal among data sets that do not clearly disagree, a claim that a hierarchical pattern of groups subordinate to groups is the universal natural order cannot be sustained as an explanandum (6, 8, 18). (That many seemingly different analyses of these data nevertheless do agree in some ways is not surprising and is discussed later.)
Tree of Life Vs Tree of cells -
Blockquote
Microbial phylogeneticists have not in general taken it to be their duty to confirm the existence of a natural inclusive hierarchy or tested the TOL hypothesis that this hierarchy is to be explained by an historical branching process. The ways in which they generally analyze and think about the molecular data presuppose a tree model, and cannot but produce trees. Even when methods that permit reticulated representations of evolution are used, the most common intent has been to discount LGT as noise, in pursuit of the legitimate “phylogenetic signal” assumed to be vertical unless significantly in conflict (11, 64, 40, 66). That is, weak signal is by default taken as vertical. Most importantly, the vast majority of analyses have consisted of the comparative evaluation of one tree with others (in search of the “true” branching topology). Seldom have investigators asked whether non-tree (reticulated) models might not better explain the data at hand. (Exceptions most often involve within-species data sets, where recombination is expected.)
Nevertheless, some phylogeneticists accept that LGT has been so pervasive over the history of life that no hierarchical classification can claim to provide the unique and true accounting of similarities and differences between organisms (and thus, an explanandum for Darwin’s explanans). Woese himself asks “What does it mean, then, to speak of an organismal genealogy when nearly all of the genes in the cell, genes that give it its general character, do not share a common history? This question goes beyond the classical Darwinian context”
The author of the paper readily accepts that we can create trees at the level of Animals. However, he denies that prokaryotes readily fall into a tree… and that modification happened in many cases by Lateral gene transfer. (Its interesting to note his field of interest is prokaryotes). This is equivalent to cutting off the trunk of the tree of life!
The final part of his argument is interesting-
Blockquote
Evolutionists have long acknowledged a diversity of population-level diversification mechanisms (selection, drift, convergence, and parallelism) and (with reservations) clade-level mechanisms that extend beyond the selectionist and gradualist framework mapped out by Darwin. At the genome level, vertical descent and LGT, gene creation, duplication and loss, in all combinations with population-level processes, expand the evolutionary repertoire. A multifaceted process pluralism is now the common view (79). The belief that nature must nevertheless exhibit a single pattern of true relationships among taxa remains vigorous, and fuels the continued enthusiasm for universal tree building and its broad application on the basis of very few and often contradictory data. We call this belief “pattern monism.” “Pattern pluralism” (the recognition that different evolutionary models and representations of relationships will be appropriate and true for different taxa or for different purposes) is an appealing alternative, and can defuse the crisis within the discipline.
Another interesting paper with respect to evolutionary trees and molecular genetics is below -
Perhaps a tree exists only is in the eyes of the beholder… (after cutting of all the “noise”)…
Again quoting interesting parts for those without time to read the paper
Blockquote
Traditionally, biologists
aspired to construct what we can call the ‘simple tree’, a ‘‘tree representing
genealogical relationships of all known species’’ (Yates et al. 2004, p. 7). This tree
is a representation of species lineages using a tree graph, that is, one composed of
recursively bifurcating branches. In the simple tree, a biological species corresponds
to a twig, and a higher clade, composed of an ancestor and all of its descendants, to
collections of branches. Every species is placed at one location on the tree, since
each species has just one parent species from which it derived.
This simple tree of life has recently been ruled ‘‘obsolete’’ (Puigbo` et al. 2009)
because it can ‘‘not provide an accurate depiction of the processes that have shaped
life’s history’’ (Fournier et al. 2009, p. 2229). Genomic evidence shows that
organisms—particularly prokaryotes—receive genes from a variety of sources,
sometimes from organisms in other species. This happens via entry of foreign DNA
into a cell followed by integration of that DNA in the host genome, in a process
called lateral gene transfer (LGT) (Planet 2002). Because of LGT, the genealogical
histories of different parts of the genome can differ and it is not possible to represent
all these different histories using a single, recursively bifurcating graph. Species
which have received genes from multiple parent species cannot be positioned on a
single branch of the simple TOL.
Blockquote
In light of this challenge to the simple tree, researchers have, broadly speaking,
taken up one of two remaining options. One option is to tailor our understanding of
the TOL so that it can accurately represent the biological facts (Galtier and Daubin
2008; Ciccarelli et al. 2006; Puigbo` et al. 2009). The tree may, for example, be
understood to represent the history of particular core genes, those which were not
transferred laterally between species. Or the ‘tree’ could be, technically speaking,
some other kind of graph, including one that allowed reticulation. Let’s call all
modifications of the simple tree ‘refined trees,’ which come in different varieties
depending on the details. After making the appropriate modifications, proponents of
this approach maintain that a Tree of Life really does exist (Lienae and DeSalle
2009; Gribaldo and Brochier 2009).
A second option is to reject the Tree of Life altogether, be it simple or refined.
Ford Doolittle and collaborators have been instrumental in arguing for this
alternative (Doolittle 1999; Doolittle and Bapteste 2007; Gogarten et al. 2002).
These researchers suggest that frequent inter-species gene transfer does not simply
obscure the tree of life, but deconstructs it. Tree-detractors may admit that refined
trees represent something—but not enough to have a legitimate claim to the
illustrious title, the Tree of Life. After all, some refined trees reflect only a tiny
quantity of genetic data, and even optimistically this may be only 1% of the total
genomic history of organisms (Dagan and Martin 2006). Detractors urge that it
would be wise to abandon this degenerating tree research program and admit that
the tree of life does not exist (Bapteste and Boucher 2008; Lopez and Bapteste 2009;
Lawton 2009).
Blockquote
A second option is to reject the Tree of Life altogether, be it simple or refined.
Ford Doolittle and collaborators have been instrumental in arguing for this
alternative (Doolittle 1999; Doolittle and Bapteste 2007; Gogarten et al. 2002).
These researchers suggest that frequent inter-species gene transfer does not simply
obscure the tree of life, but deconstructs it. Tree-detractors may admit that refined
trees represent something—but not enough to have a legitimate claim to the
illustrious title, the Tree of Life. After all, some refined trees reflect only a tiny
quantity of genetic data, and even optimistically this may be only 1% of the total
genomic history of organisms (Dagan and Martin 2006). Detractors urge that it
would be wise to abandon this degenerating tree research program and admit that
the tree of life does not exist (Bapteste and Boucher 2008; Lopez and Bapteste 2009;
Lawton 2009).
Note: Most people mean the simple tree when they refer to common decent as opposed to any tree of cells or conserved genes…
Blockquote
Yet here we see that LGT prevents the TOC from performing one task that the
simple tree could perform—explaining the hierarchical character of taxonomy.
Nevertheless, it would be overhasty to reject the refined tree just because it doesn’t
serve this particular purpose. We need to ask whether there are any other important
purposes for which the TOC might be more suited
So the question to ask is … if the so called simple tree of life proposed by Darwin doesn’t really exist… the what will happen to common ancestry… can we ever prove it or falsify it?
Prediction #1: Charles Darwin and the long tongued moth. Not discovered until 20 years after his death.
Prediction #2: From Darwin in Chapter 6 in the Origin of Species: “If it could be proved that any part of the structure of any one species had been formed for the exclusive good of another species, it would annihilate my theory, for such could not have been produced through natural selection.” - that was a successful prediction as well without any falsifications ever to date.
Prediction #3: A thousand years ago, just about every remote island on the planet had a species of flightless bird. Evolution explains this by saying that flying creatures are particularly able to establish themselves on remote islands. Some birds, living in a safe place where there is no need to make sudden escapes, will take the opportunity to give up on flying. Hence, Evolution predicts that each flightless bird species arose on the island that it was found on. So, Evolution predicts that no two islands would have the same species of flightless bird. Now that all the world’s islands have been visited, we know that this was a correct prediction. Maybe you don’t like this one but Biogeography is another field that makes no sense in light of a supernatural creation: Biogeography - Wikipedia
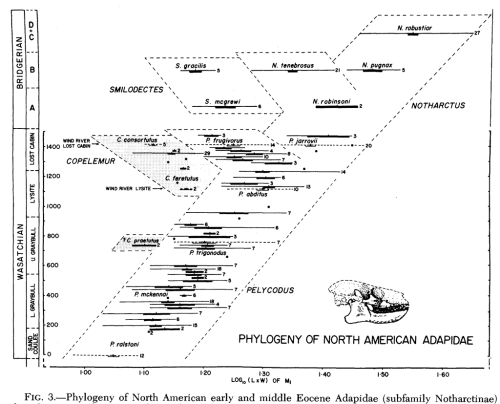
Nonsense. These giant blockquotes (can you just grab the relevant portion) and quote mines are quite tiresome. Do you know what those like Bapteste actually argue for? Or why he suggests that a ‘tree of life’ doesn’t exist? Or why this has nothing to do with falsifying let’s say common descent? The researchers are simply suggesting that inter-species gene transfer makes constructing a nice little tree much more difficult. Note this simple sequence of fossils where the y-axis represents depth of the fossils and hence age:
Is this tree ‘real?’ Well yes and no. We see the skull size following a general trend to the larger side and then we see a messy type of split-likely indicating two populations split. Are the branches drawn ‘real’ branches? Well no, but based upon such fossil sequences you can determine general relationships that ARE confirmed by genetic information with some small tweaks.
This is the last of a series but demonstrates some of the changes made to various phylogenies as we learned more data:
2 Likes
gbrooks9
(George Brooks, TE (E.volutionary T.heist OR P.rovidentialist))
188
While I spend some time reviewing your substantial text blocks… I should remind you that in prior quotes you accused ME of intellectual dishonesty and deception. Not pretty.
What I glean in reading them carefully is that biology (in general) and its major theory, evolution (in particular), are not completed works. The theory and its formulation are always being refined as biologists do their work.
So yes, the popular summaries of evolution often do not do justice to the complexities of working with the theory in real life. Some popular summaries are sloppy, and use tautological definitions. The papers you have cited are useful reminders that we have much still to learn, and much to refine even in what we have already learned.
Having cited the papers, however, you commit the non sequitur fallacy by claiming that the whole theory of evolution should be discarded. Not improved and refined, but trashed.
Actually, it is not in the least what you are saying. I draw your attention to the statement you seem to have overlooked:
Moreover, note the date of publication: 2007. Since then, Theobald et al. published a study based on a fourth kind of evidence, genomic. The only question raised about Theobald’s work is whether the nested hierarchy goes all the way back to a single first ancestor, or whether it disappears in a prokaryotic bush over a billion years ago.
Let me repeat that: none of the work you have cited disputes the ability to reliably build nested hierarchies going back hundreds of millions, if not billions, of years; and more recent genomic studies have reinforced the reliability of the nested hierarchies. The papers you cite demonstrate only that the jury is still out as to whether you can get back to a single ancestor 2 or 3 billion years ago.
So what’s the problem? In a nutshell: you seem, my friend Ashwin, inclined to leap to the unwarranted conclusion that any claim of a nested hierarchy in the animal kingdom, or among tetrapods, or among mammals, or even among primates, is just garbage non-science. But none of the evidence you have cited even begins to support this conclusion.
We can infer a more refined version of common ancestry, one in which Pan Troglodytus and H. Sapiens share a common ancestral population; in which the ape family and the canine family share a common ancestral population; in which the mammal class and reptile class share a common ancestral population; and so forth. This is the something that even the tree-detractors admit, per one of your block quotes:
I imagine that after pausing for another week or two, you will be tempted to try for a third time to find even more papers that raise the question of whether nested hierarchies can reach back to a single universal common ancestor (or not). And then you will once again be tempted to claim that the whole theory of evolution, including more modest and widely accepted corollaries such as the common ancestry of primates, is awash with hypocrisy and error. If you do succumb to the temptation, you will be wrong again.
Please prove me wrong, Ashwin. Please spare yourself and your readers the unfruitful effort of analyzing the same questions over and over again, with the only difference being a new set of papers that once again do not say what you say they say.
Surely we have better ways to redeem the time, I submit. I draw your attention to the fact that you have not yet responded to these issues
But if you prefer not to address those issues, so be it.
T_aquaticus
(The Friendly Neighborhood Atheist)
190
That is absolutely, 100% false. Phylogenies are OBJECTIVELY and EMPIRICALLY determined. They aren’t opinions.
“The degree to which a given phylogeny displays a unique, well-supported, objective nested hierarchy can be rigorously quantified. Several different statistical tests have been developed for determining whether a phylogeny has a subjective or objective nested hierarchy, or whether a given nested hierarchy could have been generated by a chance process instead of a genealogical process (Swofford 1996, p. 504). These tests measure the degree of “cladistic hierarchical structure” (also known as the “phylogenetic signal”) in a phylogeny, and phylogenies based upon true genealogical processes give high values of hierarchical structure, whereas subjective phylogenies that have only apparent hierarchical structure (like a phylogeny of cars, for example) give low values (Archie 1989; Faith and Cranston 1991; Farris 1989; Felsenstein 1985; Hillis 1991; Hillis and Huelsenbeck 1992; Huelsenbeck et al. 2001; Klassen et al. 1991).”
A web of life may very well be more appropriate if we include prokaryotes in our comparisons. Lateral genetic transfer is relatively common in prokaryotes which can mask the signal from vertical genetic transfer. However, for eukaryotes the tree of life analogy as well as phylogenies still stand.
This is a quote from some of the same authors you are referencing:
“To be sure, much of evolution has been tree-like and is captured in hierarchical classifications. Although plant speciation is often effected by reticulation (80) and radical primary and secondary symbioses lie at the base of the eukaryotes and several groups within them (81, 82), it would be perverse to claim that Darwin’s TOL hypothesis has been falsified for animals (the taxon to which he primarily addressed himself) or that it is not an appropriate model for many taxa at many levels of analysis. Birds are not bees, and animals are not plants.” Doolittle and Bapteste, 2007
So do you agree that the evidence clearly shows a tree of life for all animals and is evidence for all animals sharing a common ancestor and evolving from that common ancestor?
Did you read it? Here is a quote from that paper:
" Although this reticulation is most extensive in the evolution of prokaryotes, eukaryotes have also increasingly been caught in the act [23-26]. The comparative infrequency of HGT in the eukaryote part of the biological world means, however, that in this case the conceptual implications for the TOL might not be as drastic: the evolutionary histories of many eukaryotes appear to produce tree-like patterns (e.g., [27]). "
A question @Ashwin_s never seems to get around to answering. Even when it is pointed out to him that page 1 of the paper he is quoting doesn’t support his quote mine.
I appreciate the engagement, Ashwin. My life is in a particularly busy spell and to do justice to your response would take more reading than I’m able to commit to right now. (I can’t even particularly justify taking the time to read the response I’m currently replying to… I haven’t read all those block quotes.) I think others have already adeptly addressed the main lines of your argumentation, though. I trust that their responses are sufficient, though I’m fairly sure you won’t find them convincing.
To what extent can we “refine” the theory of evolution? This theory seems to be like theseus’ ship!
From Darwin’s times, they have pretty much refined and replaced every component of the theory.The last standing component was common descent as represented by a universal treel of life.
Now scientists are saying the said “tree of life” does not exist. That this part of Darwin’s hypothesis has been falsified.
So we come back to my first question. What exactly is the theory of evolution? Is it a falsifiable theory or some kind of unfalsifiable worldview.. a Ship of theseus that can have every component falsified and replaced but still claim to be the same old ship???
Draw the tree.. compare it to Drawins hypothesis.. place a creationists tree alongside it. If you were classifying types of trees.. The current tree accepted by consensus would be classified along with that of creationists as an unrooted tree…
Think on it.
Genomic studies do not represent what happened to particular species… it doesn’t or atleast shouldn’t generate a species tree. Biologists have arrived at trees for decades by removing evidence that pointed to a non-tree reality at the biochemical level. Once they correct this bias, and repeat their studies, we will know what the real picture is.
Now you are just repeating a statement of faith. The papers point to a methodolical error in how people have been constructing trees. they have been ignoring the major part of the genetic information. 99% in case of universal tree of life!. let me quote from a paper on eukaryote origins -
Blockquote
How we think we should depict evolutionary history has, of course, an enormous effect on how we analyse data, how we write analytical software and how we depict the final results. If we feel that the evolutionary history of a dataset has been tree-like, then it is likely that the first, perhaps only, analyses we carry out will be a phylogenetic analysis using software that generates, as an output, a tree. We know of course that human populations do not have a tree-like history; therefore, we are usually disinclined to use software for tree reconstruction to depict these histories [2]. A phylogenetic tree can always be derived based on the complete genomes of two parents and their children. However, we know that the tree will be meaningless, because a tree-like process did not generate the data. What this means is that the outcome of an evolutionary analysis is always contingent on our a priori opinions for how the data have evolved. In some cases, as in the above example, our knowledge of the process that generated the data is good enough to let us unambiguously avoid the use of a particularly poorly fitting model (i.e. a tree) to describe the data (i.e. the relationships between the genomes of two parents and their progeny). However, in most cases, we lack the knowledge to unambiguously reject a model (or a class of models) based on previous observations. In such cases, a better course of action would be to consider a variety of models and ask which fits the data best (if not adequately).
For now we can say that. But since all our conclusions are based on analyses that assumed trees, we cannot be sure whether it actually is what the data* says.. Its an interpretation forced by an a priori assumption.Maybe the interpretation is true and agrees with the data.. maybe not..
However, even of higher phyla show tree like patterns, what you will get is an unrooted tree..Like creationists draw. Not the kind predicted by the modern synthesis.
An unrooted tree is closer to what someone like Behe, or any creationist (or someone like shapiro) would predict rather than what the modern synthesis does.
Its probable that early eukaryotes also do not really show any tree-like organisation (see paper below).
The paper describes the eukaryotes not arising from any particular group (i.e archaebacteria and eubacteria), but rather as a mix and match between the two from some kind of “symbiotic” event. i.e its something of a chimera. (not descent via modification). http://rstb.royalsocietypublishing.org/content/370/1678/20140323
In short, what we have is not common descent. What we have is two or possibly three unrooted trees.
And the fact that the analysis of all trees we have is based on a biased interpretation of data.
I will come to it when i have time (mainly because i am curious). I have put it on the back burner because its will not prove anything. if it turns out to be diversity in species or kinds.. you will still claim its vestigial.
Even if it turns out to be vestigial, it will still not make any difference. because, i have already accepted in principle that some ammount of adaptation does happen. (Even YEC’s accept that). It seems to be an excercise in futility and perhaps a never ending chain.
I accept that the authors do not question the tree like nature of animal classification. I have noted in one of my posts that their main area of expertise/work is not animal classification. I will come to animal classification later on.The main points i wish to make from the papers are as below -
The so called tree of life is unrooted.. i.e it does not end ins LUCA. It is actually three separate trees.. (as of now).. and perhaps more appropriately 3 separate networks of genetic relationships which do not necessarily show descent.
Observations of a tree like appearance can be because of assumptions of a tree.
Would you agree to the same?
The authors make a case that looking for tree in Biological data is a wrong method and can influence programs to give skewed data. hence they propose that Scientists should look for networks of relationships to get unbiased data from which to draw conclusions.
Some people have carried forward such a research programe.. And perhaps it will turn out that genomes of animals show a nested heirarchy… perhaps not. That is yet to be determined through empirical science.Though i would expect to find trees at some point.. either among animals.. or mammals.. or atleast families… (But how would that differ from my Automobiles example??)
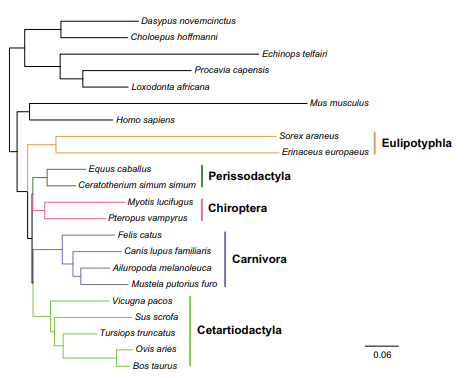
Below is an interesting study within mammals.
Blockquote
With the advent of more genome data becoming available, along with the ability to explore deep divergences in greater detail, it is becoming evident that evolutionary processes are best interpreted as networks. Networks naturally highlight the conflict and difficulties of previous phylogenetic studies to find a congruent bifurcating tree within this group. The hope of phylogeneticists that whole genome data would one day yield a single, stable and bifurcating evolutionary tree [18], [63], [64], is not fulfilled for some parts of the placental mammalian tree. However, it seems that a more valuable lesson can be learned from genome analyses. That is, some divergences are not characterized by bifurcations but rather that the evolution of some placental mammals represent a complex pattern of genealogies of different parts of the genome. Speciation processes that can be revealed from genome data even for deep divergences, define this pattern. The evolution of Carnivora, Perissodactyla, Chiroptera, and Cetartiodactyla (Laurasiatheria) represent such a case.
The bare minimum we can conclude from the paper is that Genome studies do not support a bifurcating tree of life for Laurasiatherian mammals. The data better supports a network.
A tree has to be force fitted.
I dont expect network analysis to be very popular when studying animals.. because we human beings think more easily in terms of nested heirarchies.
So give it a decade or two to catch on..
Think.. what exactly is an “interspecies gene transfer”?
Bapteste is quite clear.. he emphatically says that Darwins hypothesis of the tree of life has been falsified… Can he be any more clear???
I gave extensive quotes to avoid being accused of “quote mining”.. so that the context is clear. let me just quote the relevant part as you have requested -
Blockquote
Darwin claimed that a unique inclusively hierarchical pattern of relationships between all organisms based on their similarities and differences [the Tree of Life (TOL)] was a fact of nature, for which evolution, and in particular a branching process of descent with modification, was the explanation. However, there is no independent evidence that the natural order is an inclusive hierarchy, and incorporation of prokaryotes into the TOL is especially problematic. The only data sets from which we might construct a universal hierarchy including prokaryotes, the sequences of genes, often disagree and can seldom be proven to agree. Hierarchical structure can always be imposed on or extracted from such data sets by algorithms designed to do so, but at its base the universal TOL rests on an unproven assumption about pattern that, given what we know about process, is unlikely to be broadly true.
Note: Bapteste who woeks mainly on prokaryotes is emphatic that prokaryotes dont fall into a tree pattern. He believes animals do..
However, that may not be the case. as suggested by a paper i have referred to above..
Even if animals do fall into trees, we have unrooted trees lwhich is closer to a creationist expectation that Darwin’s.
Sorry if i gave the wrong impression… I don’t believe you intend to deceive any one.My impression is that you are genuinely sincere.
However its possible for anyone of us to fall for strategies in discussions which are inherently deceptive..
gbrooks9
(George Brooks, TE (E.volutionary T.heist OR P.rovidentialist))
195
And yet … you call one of the BioLogos Mission Statements as deceptive - - when in fact it goes to great lengths to avoid any deception! To paraphrase:
o Evolution on Earth requires God;
o Any model of Evolution of that excludes God’s participation is rejected;
o Evolution must include a teleological role for God.
So, what you are saying is that your own position is also deceptive? If it isn’t, how exactly do your positions differ from the positions set out by BioLogos?
T_aquaticus
(The Friendly Neighborhood Atheist)
196
Given that you don’t accept the refined theory, does it really matter?
The one mechanism that Darwin put forward was natural selection, and it is still a central pillar of the theory.
“To be sure, much of evolution has been tree-like and is captured in hierarchical classifications. Although plant speciation is often effected by reticulation (80) and radical primary and secondary symbioses lie at the base of the eukaryotes and several groups within them (81, 82), it would be perverse to claim that Darwin’s TOL hypothesis has been falsified for animals (the taxon to which he primarily addressed himself) or that it is not an appropriate model for many taxa at many levels of analysis. Birds are not bees, and animals are not plants.”
Doolittle and Bapteste, 2007
Want to try that one again?
What creationists are claiming that all eukaryotes share a common ancestor and evolved from that common ancestor?
The tree is real for very large groups of species, such as the eukaryotes. Do you accept common ancestry for all eukaryotes? If not, then why make the argument you are trying to make?
The tree for animals has already been determined empirically, as the authors of the papers you are quoting have already pointed out.
It seems to me that you are willing to agree with the biologists that descent with modification explains from the scientific perspective how the primates are descended from a common ancestral population, and that descent with modification, be it structured as a tree or as a network, explains from a scientific perspective the origin of species both small and large. You do have serious reservations about origin-of-life pronouncements, and think that the first cellular organism was the result of God’s direct intervention–i.e., a miracle–rather than through regular scientifically tractable means. You also believe that any tree that biologists can reconstruct will only reach back to the origin of multi-cellular life some 630 MYA.
If I have understood you correctly, we are largely in agreement on these points. The only substantial difference I see between us is that I am open to the possibility OOL researchers may be able, someday far in the future, to infer a scientifically tractable mechanism through which biological life on earth came to exist. Should that day arrive, I would regard the mechanism as an exhibition of God’s marvelous creativity and design. For today, however, I would agree that the RNA world is a hypothesis rather than a theory.
One of the most recent sources you cited in another thread reached this conclusion:
I agree with this conclusion that Marvin quoted. If you are willing to agree with that conclusion, too, then we can throw a party, hand out trophies, and move on to other pursuits.
Grace and peace,
Chris
3 Likes
T_aquaticus
(The Friendly Neighborhood Atheist)
198
So do you agree with the authors?
“We exemplify this on a subset of 1159 suitable genes that have individual histories, most likely due to incomplete lineage sorting or introgression, processes that can make the genealogy of mammalian genomes complex.”
It’s not as if some of the genes they looked at demonstrated more relatedness to jellyfish or amphibians for some mammal species. All of the genes they looked at put these species well within the mammal clade. The only arguments they are putting forward is for the fine details of that tree, and the noise within that tree caused by very real biological processes like incomplete lineage sorting. It is an argument about noise being high when looking at the deep nodes of a tree WHICH IS EXPECTED WITH EVOLUTION. You might as well claim that a blurry photo of a distant galaxy means that galaxies don’t exist.
“To be sure, much of evolution has been tree-like and is captured in hierarchical classifications. Although plant speciation is often effected by reticulation (80) and radical primary and secondary symbioses lie at the base of the eukaryotes and several groups within them (81, 82), it would be perverse to claim that Darwin’s TOL hypothesis has been falsified for animals (the taxon to which he primarily addressed himself) or that it is not an appropriate model for many taxa at many levels of analysis. Birds are not bees, and animals are not plants.”
Doolittle and Bapteste, 2007
You are basing your conclusions on a 2011 paper. However, later research that used a much larger dataset strongly refuted the conclusions of that paper. The 2017 paper is:
Which means Biologos rejects the Scientific theory of evolution which is a theory that excludes Gods participation and excludes a teleological role for God.
My position is not deceptive because i don’t use the term evolution to describe a belief that is patently not Evolution and in fact denies evolution is ultimately a natural process.
Yes it does. It shows whether the so called “theory” is falsifiable .. and what exactly the theory itself is. For example, if evolution as a theory can be refined ad infitum and all its components replced at some point or the other, its not a falsifiable theory. Its an over arching philosophy which is essentially untestable.
I hold that the idea of evolution is unfalsifiable.. and every component which can be falsified has been falsified.
Ever heard of the neutral theory of evolution? Natural selection has been cut down to size. It is not what Darwin claimed it would be… i.e a unifying principle. Its valid in some cases/example of evolution and invalid in others..
Its also untestable/unfalsifiable.. Biologists are even now looking for a definition of Natural selection that would permit falsification.
Yes.. the point being that Tree of Life is Falsified.. At best.. its a “Tree of animals”…
not even plants and animals. Reticulated evolution is another word for data that is not tree like.. it refers to a network.
You have lost Prokaryotes, eukaryotes and plants…
Even creationists wouldnt have a problem with the last comment
** Birds are not bees, and animals are not plants.”**
Even Biologist are not claiming eukaryotes have a common ancestor.. The current understanding is that eukaryotes are a hybrid of Eubacteria and archebacteria traits/genes.. also cyanobacteria contribute to plants at some level too..
I dont think anyone even claims the first hybrid gave rise all the families of eukaryotes.. thats also a reticulated mess and scientists are trying very hard to fit it into a tree..
Why would creationists make such a claim when even evolutionary biologists hesistate to?
My point is that Biologists are now beginning to repesent life as creationists wouls… as a unrooted network on an overall level with trees found among more similar organisms.
No the idea is, life is not a tree.. its a network of gene sharing.
Are you claiming that common ancestry started only with eukaryotes?
And if most unicellular eukaryotes turn out not to show tree like patterns (which is a highly likey scenario considering the various conflicting results with respect to where the eukaryote Ca is positioned in the tree), would you claim common ancestry started out only with multicellular organisms? And what do you do when plants show extensive reticulation (as they are shown to do)?
Yet they somehow feel compelled to propose a network type of relationship.. reticulated networks are currently being used to show the relationship between birds also for your information.. Basically we have a broken chain of several tree like patterns. this is not the unbroken chain of descent predicted by Common ancestry.
I was pointing out where science stands currently in my view.I am currenlty agnostic on primate evolution.I am not philosophically commited to any from of scientism as it stands on a false belief in the inerrance of scientific consensus.Let me give an example. In Einsteins times, it was the opinion of most scientists including Einstein that the universe was eternal, and went through and endless cycle of expansion and collapse. If any one changed their theology based on Einsteins then scientifically valid view, they would have been in great error.Though i do read science, i am very careful about what i believe is truth with a capital T. I allow philosophy and world views (including views on the bible) to educate my final beliefs and i would expect this from any rational person.
I prefer a wait and watch policy with respect to scientific consensus. My theological views are formed in the traditional way. reading the bible and praying.
I expect the next few decades to show the following based on my understanding of the issues-
That natural genetic engineering is responsible for novelty/and big changes in phenotypes/morphology.(LGT, endo-Symbiosis, Hybridisation, and other yet to e defined processes that achieve phenotypic change through inserting pre-defined genetic content into a host genome).
As knowledge of complete genomes increase, i expect reticulated networks to emerge as a better representation of the relationships among species even in more complex domains such as the animal kingdom.
That someone would redefine evolution to mean “Descent with modification”, where said modification is not restricted to causes related to lineage or something along those lines..
Textbooks will be updated decades after the fact with new just so stories..
There is a big need for a paradigm shift in biology..
Its possible to arrive at a statistical tree of life (i.e a tree of trees) and also a tree of life from single genes. I don’t dispute that. However, there is no reason why one set of genes should tell a different story from the other if both are inherited from a CA. Its the overall picture that is reticulated.. I dont think the 2011 paper disputes that Trees of life can be created..
They just tell different stories based on the genes you select. Introns tell different stories.. coding sequences tell another story.. micro Rna based comparisons tell something else.
If the introns are inherited from a CA, then where do the CDS and other gene sequences which tell a different story come from?
This is why Scientists like Baptiste say that these genomic trees need not really represent the history of a species vis a vis its lineage.Its just a way to classify or represent organisms.