Hi Steve,
See: Index of /goldenpath/hg38/vsPanTro6 and Index of /goldenpath/hg38/vsPanTro4
Here is the information available at the first URL above:
This directory contains alignments of the following assemblies:
-
target/reference: Human
(hg38, Dec. 2013 (GRCh38/hg38),
GRCh38 Genome Reference Consortium Human Reference 38 (GCA_000001405.15))
-
query: Chimp
(panTro6, Jan. 2018 (Clint_PTRv2/panTro6),
University of Washington)
Files included in this directory:
-
md5sum.txt: md5sum checksums for the files in this directory
-
hg38.panTro6.all.chain.gz: chained lastz alignments. The chain format is
described in Genome Browser Chain Format .
-
hg38.panTro6.net.gz: “net” file that describes rearrangements between
the species and the best Chimp match to any part of the
Human genome. The net format is described in
Genome Browser Net Format .
-
hg38.panTro6.net.axt.gz: chained and netted alignments,
i.e. the best chains in the Human genome, with gaps in the best
chains filled in by next-best chains where possible. The axt format is
described in Genome Browser axt Alignment Format .
-
hg38.panTro6.synNet.maf.gz - filtered net file for syntenic alignments
only, in MAF format, see also, description of MAF format:
Genome Browser FAQ
-
hg38.panTro6.syn.net.gz - filtered net file for syntenic alignments only
-
reciprocalBest/ directory, contains reciprocal-best netted chains
for hg38-panTro6
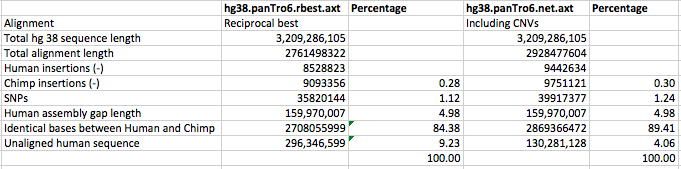
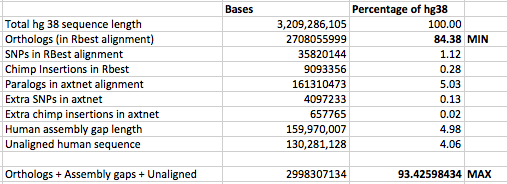
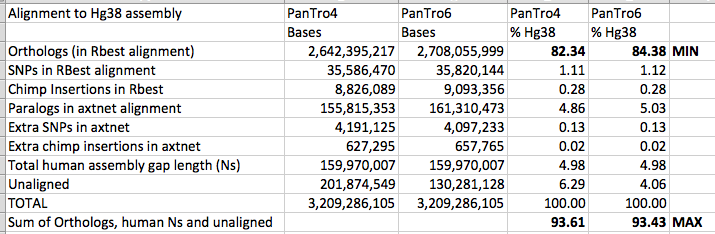
The hg38 and panTro6 assemblies were aligned by the lastz alignment
program, which is available from Webb Miller’s lab at Penn State
University (CCGB: Miller Lab). Any hg38 sequences larger
than 20,010,000 bases were split into chunks of 20,010,000 bases overlapping
by 10,000 bases for alignment. A similar process was followed for panTro6,
with chunks of 20,000,000 overlapping by 0. Following alignment, the
coordinates of the chunk alignments were corrected by the
blastz-normalizeLav script written by Scott Schwartz of Penn State.
The lastz scoring matrix (Q parameter) used was:
A C G T
A 90 -330 -236 -356
C -330 100 -318 -236
G -236 -318 100 -330
T -356 -236 -330 90
with a gap open penalty of O=600 and a gap extension penalty of E=150.
The minimum score for an alignment to be kept was K=4500 for the first pass
and L=4500 for the second pass, which restricted the search space to the
regions between two alignments found in the first pass. The minimum
score for alignments to be interpolated between was H=2000. Other blastz
parameters specifically set for this species pair:
E=150
M=254
O=600
T=2
Y=15000
The .lav format lastz output was translated to the .psl format with
lavToPsl, then chained by the axtChain program.
Chain minimum score: 5000, and linearGap matrix of (medium):
tableSize 11
smallSize 111
position 1 2 3 11 111 2111 12111 32111 72111 152111 252111
qGap 350 425 450 600 900 2900 22900 57900 117900 217900 317900
tGap 350 425 450 600 900 2900 22900 57900 117900 217900 317900
bothGap 750 825 850 1000 1300 3300 23300 58300 118300 218300 318300
Chained alignments were processed into nets by the chainNet, netSyntenic,
and netClass programs.
Best-chain alignments in axt format were extracted by the netToAxt program.
All programs run after lastz were written by Jim Kent at UCSC.
References
Chiaromonte F, Yap VB, Miller W. Scoring pairwise genomic sequence
alignments. Pac Symp Biocomput. 2002:115-26.
Kent WJ, Baertsch R, Hinrichs A, Miller W, Haussler D.
Evolution’s cauldron: Duplication, deletion, and rearrangement in the
mouse and human genomes. Proc Natl Acad Sci U S A. 2003 Sep
30;100(20):11484-9.
Schwartz S, Kent WJ, Smit A, Zhang Z, Baertsch R, Hardison RC,
Haussler D, Miller W. Human-Mouse Alignments with BLASTZ. Genome
Res. 2003 Jan;13(1):103-7.
{kind=link}