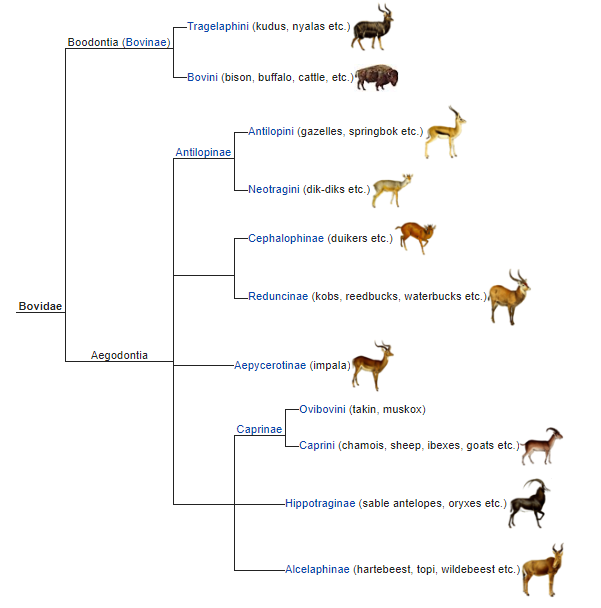
Sheep actually make for a really very interesting case study, since you mention it.
Have you ever looked at a family tree of the clade Bovidae?
(Source:
Wikipedia.)
In English, we consider sheep and goats as quite distinct from cows! But there are places you can go in the world where the cows are quite small and the sheep are quite large, and the sheep’s neck is even generally in a straight line with his spine, and you look at them and say, “Wow, you know, cows and sheep really do look quite similar.” In fact, the entire Bovidae family tree looks quite similar, and it’s not hard for me to imagine them all diverging from a common ancestor over the past 20 million years, as asserted by evolutionary theory. So… was this micro- or macro-evolution?
For comparison’s sake: In one language in sub-Saharan Africa, there is no one catch term for hyenas. The word for spotted hyena is îgr̰im or dondo (depending on the dialect) and the word for striped hyena is doro or daro (depending on the dialect).
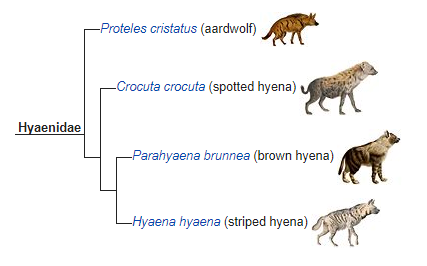
(Source: Wikipedia.)
The hyena family also originated about 20 million years ago and has also diverged, but in English we call them all (except the aardwolf) “hyenas” and most of us Anglophones would have no problem saying that they’re basically the same “kind,” if you want to go the baraminology route.
Does what we consider microevolution versus macroevolution depend on our cultural baggage? Or can we honestly say that we know a “kind” when you see one? Can you really not imagine, over 20 million years — that’s about 10,000,000 generations, give or take — that a sheep could become a goat or a cow or an antelope?
[Edit… Having now read the rest of the thread, I see that most of these points have already been made, and Chris has admitted that the line between micro- and macro- is somewhat fuzzy. But I’ll leave my post up anyway in case it’s of interest to anyone (and because I spent a lot of time on it, haha).]