I would hope that you have better things to do than waiting for my posts.

A better analogy to evolution would be that you have two editions of OtOoS, in which case you’d have more information than you would with just one, but not twice as much.
1 Like
@aarceng (& @Lynn_Munter and @T_aquaticus):
I neglected to point out two things… my life has been a bit hectic as of late…
-
I assume we can all accept adding the small bit that traditionally goes with Macro-Evolution:
the part that conventionally accommodates the idea that Macro-Evolution refers to the kind of evolution that creates a new species (with the usual YEC proviso - - assuming that the emergence of a new species is even possible. -
The more interesting fine point is one that was described to me by some sort of angst-ridden deist. He quoted Ernst Mayr, the grand old man of modern Evolutionary Science… and the father of the Genetic Definition of Species/Speciation:
“In his book [[Systematics and the Origin of Species (1942)]] he wrote that a species is not just a group of morphologically similar individuals, but a group that can breed only among themselves, excluding all others.”
"His theory of Peripatric Speciation (a more precise form of Allopatric Speciation which he advanced), based on his work on birds, is still considered a leading mode of speciation. . . "
“Mayr is sometimes credited with inventing modern philosophy of biology, particularly the part related to evolutionary biology, which he distinguished from physics due to its introduction of (natural) history into science.”
My source said that Ernst Mayr can be quoted as saying that “speciation” is something that occurs at the MICRO-evolutionary level. I immediately perceived what it was Mayr was saying (assuming my source’s quote is correct) . . . but I could not get agreement!
THE POINT: The point of the paragraph is that my source insisted that Mayr says Speciation is a MICRO- event, not a macro event. But misunderstood why that is so! He was trying to say that MICRO-Evolution is where we find SPECIES! All I could think was “OYYYYY”
Speciation very rarely happens in a single instant … by means of a single generation. And when it does, it is usually something associated with plant replication issues.
Speciation is a process … sometimes a lengthy one… and sometimes an interrupted one, where two groups that were seemingly on their way to becoming two separate species encounter yet another change in circumstances… and the process of speciation is halted or reversed. It is at the conclusion of this extended period of “ALMOST A NEW SPECIES” that “speciation” is something experienced WITHIN a species. Semantically speaking, it would be definitionally impossible for speciation to be experienced (initially) any other way.
Once “speciation” actually occurs, “speciation” might be said to have also ended at the very same time.
And here is a posting I did long ago… where I show that the “Kinds” of Genesis are actually consistent with Mayr’s definitions!
@@@@@@@@@@@@@@@@@@@@@@@@@
Link on Historical Background
1 Like
There is a nuance that is missing in that definition of species. A species is a population that interbreeds, or “total genetic variability of nature organized in the form of discrete packages” as described by Mayr here. It is entirely possible that two species could interbreed, but if they don’t interbreed when given the chance then they are separate species. I would replace “can’t interbreed” with “don’t interbreed”. Also from the link above:
“It is thus well established that a leakage of genes occurs among many good “reproductively isolated” species. This induced me to revise the definition of isolating mechanisms to “biological properties of individuals which prevent the interbreeding [fusion] of populations”(1970, 56). Thus, isolating mechanisms do not always prevent the occasional interbreeding of non-conspecific individuals, but they nevertheless prevent the complete fusion of such species populations.”–Mayr
As to micro/macro evolution, I found this on wiki:
“transspecific evolution is nothing but an extrapolation and magnification of the events that take place within populations and species…it is misleading to make a distinction between the causes of micro- and macroevolution”.–Mayr
I would not personally include this in my definition, although you are welcome to in yours. The whole question of where exactly speciation is seems overly complex. I don’t see a point in trying to draw a hard line through it to divide micro from macro. In fact, from an evolutionary perspective such a line cannot be drawn, so I am perfectly comfortable leaving my definitions with the vague “relatively large” and “relatively small.”
But I agree with your post about ‘kinds!’ Nothing indicates immutability.
Well, yes, but sometimes they’re boring and I like to procrastinate. ![]()
1 Like
I can understand why you are attracted to the “nuance” of this. It helps bridge the idea of “Species” between the old phenotypical definitions and a strict reproductive one… and so spares headaches in that regard. But I think for the purpose of Biblical Metaphysics … this popular solution is not crisp enough.
I read through the quotes and references for Mayr in your link. And I don’t see Mayr actually embracing this “more nuanced” idea, as attractive as it can be in some circumstances. Naturally, if you think there IS a Mayr embrace of this definition (him preferring the concept of “they don’t” instead of “they can’t”), I do want to have this firmly and correctly embedded in my brain. So please do correct me with the quote that would do so. And I will faithfully reproduce that evidence whenever needed.
Short of such correction, however, let me offer you my approach to the “Biological Concept of Species” (aka the oft-mentioned BSC from the article):
Every elementary-aged child “knows” that Lions and Tigers are different species. And yet they are able to successfully interbreed (and we should be careful to specify that “successful” means the next generation is also fertile). The reason we usually don’t have to discuss this oddity is that back in the common ancestry of Lions and Tigers, two populations specialized their adaptations to two different ecologies: the Tiger-ide population moved into the jungles (usually India/Asia), while the Lion-ide population came to rule the drier savannas of Africa. So… it would be be a peculiarly unhappy village that would be at the functional intersection of both Lions and Tigers!
A recent article does a great job of exploring the genetic vs. non-genetic evidence for the major branches of the Felidae. Here are a few pages (with URL and article title embedded):
[ Be sure to click on images for maximum legibility ]

[ Be sure to click on images for maximum legibility ]
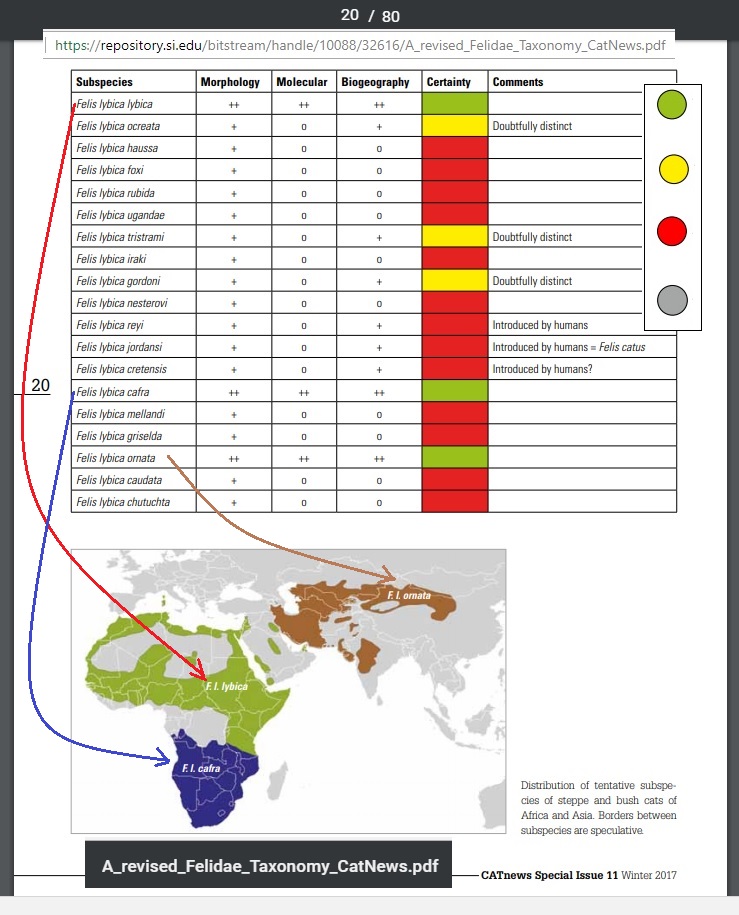
[ Pages 5 and 20 featured above! “A Revised Felidae Taxonomy” as PDF ]
^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^
This 2017 report should be cited as:
Kitchener A. C., Breitenmoser-Würsten Ch., et al.
(Eizirik E., Gentry A., Werdelin L., Wilting A., Yamaguchi N., Abramov A. V., Christiansen P., Driscoll C., Duckworth J. W., Johnson W., Luo S.-J., Meijaard E., O’Donoghue P., Sanderson J., Seymour K., Bruford M., Groves C., Hoffmann M., Nowell K., Timmons Z. & Tobe S.) 2017.
A revised taxonomy of the Felidae.
“The final report of the Cat Classification Task Force of the IUCN/SSC Cat Specialist Group.”
Cat News Special Issue 11, 80 pp.
^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^
What we see illustrated in the last two pages is a rack of RED “traffic lights” and a few yellows indicating that the non-genetic information originally used to distinguish multiple strains of this feline family are probably spurious. And the three most likely “species” are displayed on the world map. I was delighted with this example because it showed these groupings seem to correlate well with climate.
And yet, as I’ve read in a number of articles, Felines in general seem to have quite a close genetic affinity. This is probably not unique in the animal kingdom… but it is probably not what the average person expects when he is thinking LION is one “kind” and TIGER is yet another “kind”.
I would suggest that for the purpose of discussing Genesis and Biblical notions of kind, it would seem that Lions and Tigers are probably best considered a single KIND.
Genesis 1:24-25 reads: “And God said, Let the earth bring forth the living creature after his kind, cattle, and creeping thing, and beast of the earth after his kind: and it was so. And God made the beast of the earth after his kind, and cattle after their kind, and every thing that creepeth upon the earth after his kind…”
I have posted on this idea a few times already: it would seem the Bible is using the BSC concept. If a group of animals can reproduce a new (and fertile) generation - - that is a KIND! And if they cannot, they are two kinds present (or multiple kinds). Sure, it flies in the face of tradition and convention from a human viewpoint. But it is quite precise and non-arbitrary in my view… or at least less arbitrary than any other definition I’ve looked at.
The important point to make about Genesis 1:24-25 is that there is nothing in the wording that says kinds can’t biologically drift! And this is something most Creationists (even ID Creationists) don’t seem to acknowledge. They are inclined to assert that God “instantiates” specific “KINDS” … and thus the genome of the original “KINDS” is quite specific - - divinely defined even.
But does God lose interest in this precision once the populations are coping with survival and producing the next generations? I don’t think so. If, from the Christian viewpoint, we want to say that God created all the specific Kinds … I don’t think we ALSO mean that God either won’t allow “micro-evolution” to occur (aka, adaptation to their changing surroundings). And I also don’t think God doesn’t care about each Kind’s genome 4000 years later! I think it is reasonable to presume that God is just as precise with arranging for the evolution of a kind’s genome in a Creationist context as he is in an Evolutionist context!
Let me state that again - but in a slightly different way: It is reasonable to think that if God has specific ideas of what the genome is for a “kind” when he creates it (by miraculous “POOF!”)… that God is just as specific about how the population genome of a given kind CHANGES from generation to generation (just as if all creation was originally via Evolutionary processes under God’s control!).
Naturally, in the field of science, all of these metaphysical speculations are rather beside the point. And so I don’t think it is surprising, Mr. T, that you (representing one of the better classes of academic Atheists) prefer a less rigid genetic approach to defining Species. But I don’t find that approach to be a very good fit to Christian supporters of Evolution, especially if the Christian is leaning towards a “Dual/Combination” Approach for Creation!:
where some creation is via miraculous “poofing”, and other phases of creation are via naturalistic (but God-guided) Evolutionary processes. This combination approach would be generally more compatible with Old Earth scenarios.
I look forward to your thoughts, @T_aquaticus .
I have already given my reasons for not using speciation as the dividing line. YECs have no problem with speciation as such since It allows the development of many species from a relatively few Kinds on Noah’s Ark. I could see this happening by each new species having a subset of the genetic information of the Ark Kind and hence it would be speciation by information loss which I would see as incompatible with the concept of macroevolution. (Again for reasons given above.)
Tying it to speciation would also rule out macroevolution occurring below the species level. I think @sfmatheson at #154 was suggesting that adult lactose tolerance was an increase in information; macroevolution? Was the insertion mutation that produced the black Peppered Moth macroevolution? Not if it requires speciation.
I just found these comments on Uncommon Descent. Posted for information.
[12] Macro-evolution is nothing but lots and lots of “micro-evolution”!]((Frequently raised but weak arguments against Intelligent Design – Uncommon Descent)
[13] Real Scientists Do Not Use Terms Like Microevolution or Macroevolution](Frequently raised but weak arguments against Intelligent Design – Uncommon Descent)
The variants I was referring to (here) are SNPs. “Increase in information” is not how anyone would describe them. They are tiny changes in enhancer DNA sequences that cause changes in expression timing.
I thought we had already discussed the ways that evolutionary biologists use the term ‘macroevolution.’ It is rare (for them to use it), clear when it is, and almost completely unrelated to the way it is being used in this typical creationist-oriented conversation.
4 Likes
My position is that there is no dividing line you can draw anywhere. No attempt to map out original Kinds will ever conclusively be able to say, ‘these animals are related and these are not’ because there’s no natural division there for them to identify. The concept is not based in reality.
I read the thoughts at Uncommon Descent, and it fails to provide an effective, functional definition for ‘function,’ (is skunk spray a function? Is hair color a function?) That entire first response is poorly reasoned baloney based on several false assumptions. Evolutionary algorithms can in fact assemble working computer programs from random simple lines and new fuctional proteins are not very difficult to assemble randomly either, as discussed earlier. Nothing about the process of evolution prohibits multi-stage, complex processes evolving.
I am also left wondering (again) what exactly is meant by ‘body plan.’ Is each Kind meant to have its own body plan? Or are we just referring to the Cambrian explosion where we have body plans like ‘vertebrate’? Or something in between? Is ‘tetrapod’ a body plan?
I think their argument relies on a sort of armchair pontificating where nobody is actually examining the details. It might sound fine, so long as you don’t get up out of that armchair and start trying to apply it to the real world.
4 Likes
Some Creationists love to dismiss “so-called speciation” by saying what’s so special about the examples if you still have a brightly colored fish that is still a brightly colored fish?
Where is the Fish to Amphibian example, evolved right before human eyes?
Where is the Lizard to Bird laboratory study?
Where is the mammal to whale evolution witnessed in modern science?
Even the fast-motion video done by Harvard Medical School with a giant table-sized plate of agar is ho-hummed … because despite proof of “micro-evolution”, at the end of the study, you only have bacteria, not a colony of cells in the shape of Elvis.
The closest I could come to satisfying this peculiar form of Creationist melodrama are the three “Creature Plans” found in Australia … all different ecological niches, appearances and methods of living, and yet all peculiarly closely related!
I have reproduced parts of another post in another thread … pointing out that when one particular population of Marsupial made it to Australia, they discovered a continent with no rivals… no Placental Mammals… and apparently not even an earlier marsupial arrival.
And so the “diaspora” began… with each new Viking Marsupial discovering unexploited ecological niches!
In short, it is the perfect recipe for analysis: ONE KIND differentiates into at least THREE KINDS:
vegetarian mole, carnivorous canine like predator, and an omnivorous animal of the “bandicoot” phenotype!
@aarceng, the text below is for you!
@@@@@@@@@@@@@@@@@@@@@@@@@@@@@@@@@@@@
How about THIS article below! … which even if you are skeptical about the key placental link from South America to Australia, the article compares three very diverse “kinds” of Australia’s fauna and concludes that they are unusually closely related, genetically, despite the distinct “kinds” of animals they represent:
- a vegetarian mole;
- a non-jumping bandycoot type (omnivore); and
- a carnivorous hunter type (Tazmanian Devil).
^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^^
This post is just to provide the separate link of the study mentioned above:
“Tracking Marsupial Evolution Using Archaic Genomic Retroposon Insertions” by Maria A. Nilsson, Gennady Churakov, Mirjam Sommer, Ngoc Van Tran, Anja Zemann, Jürgen Brosius, and Jürgen Schmitz
PLoS Biol. 2010 Jul; 8(7): e1000436. Published online 2010 Jul 27. PMCID: PMC2910653 PMID: 20668664
Notice in the image below, the various branches associated only with South America, and branches associated with Australia, where “radiating” speciation continued, in isolation from the rest of the world.
[Be sure to click on the images to enlarge text to a more convenient font size!]
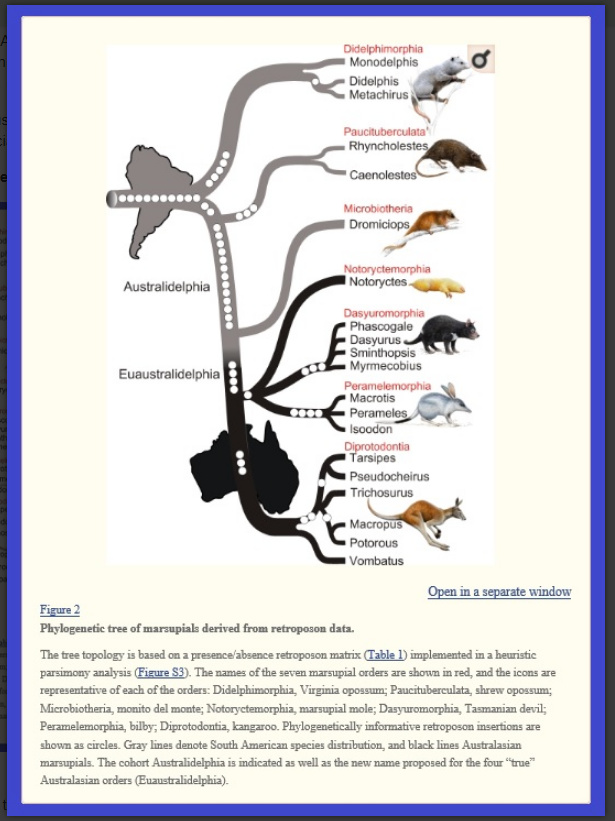
.
.
This image will be of particular value to us later on, because it creates a convenient grouping of some fairly disparate phenotypes:
While at the top we have “shrew-like” forms, and at the bottom we have “kanga” forms aggregated, in the middle grouping, we have the suggestion that three very distinct groupings share a close heritage:
Dasyuromorphia: the group having most of Australia’s carnivorous marsupials, including
quolls,
dunnarts,
the numbat,
the Tasmanian devil,
and the thylacine.
[In Australia, the exceptions include the marsupial moles and the omnivorous bandicoots.]
Notoryctemorphia: moles, vegetarian
Peremelamorphia: bandicoots & bilbies “the characteristic bandicoot shape: a plump, arch-backed body with a long, delicately tapering snout, very large upright ears, relatively long, thin legs, and a thin tail. Their size varies from about 140 grams up to 4 kilograms, but most species are about one kilogram, or the weight of a half-grown kitten [4 kilograms = 4 half-grown kittens].”
.
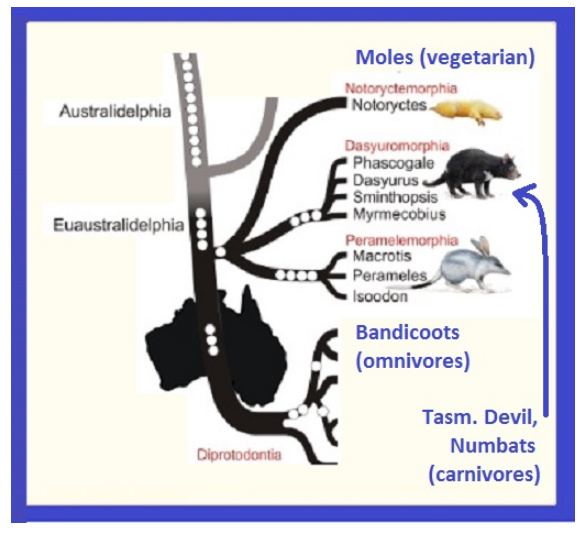
.
This is the ideal “research scenario” to see how much genetic change occurs, and how quickly - - according to Evolutionary Theory - - to accomplish divergence into three distinctive “forms” of marsupials!
.
.
2 Likes
So, two cats?
Catchy visual. I usually visualize a kilo as a 1000 cc of water, or about a quart jug of milk. Sort of a smallish cat.
But, really enjoyed the post, George. Great example.
1 Like
Frankly, @Lynn_Munter, I was so busy laughing at the writer’s use of a “Half Grown Kitten” as a frame of reference, I couldn’t help offering an equally obtuse reference, by multiplying it by four!
Just a few minutes ago I found this website:
https://thewebsiteofeverything.com/animals/mammals/adult-weight.html
which was surprisingly unhelpful. I suppose mostly because there were just too many choices to make at the 1 kilo level of existence!
Animals that as adults are about 4 kilos include the White-tailed mongoose! But I certainly have no first hand experience as to how big that is in real life. Are mongoose (plural?: mongeese?) without white tails very much different? I’m in no position to say!
But it did say an adult skunk is about 4 kilograms. Now SKUNKS I know!
With this encouragement, I moved back to the 1 kilo part of the list and was immediately hit with the Coppery Titi. I had no idea that the Coppery Titi was so light… I suppose because I thought they were imaginary creatures!
The Black Flying Fox is ALSO 1 pound … but can I really take that seriously? One: it isn’t a fox. Two: if it was a fox, it would not really be flying. A one-pound BAT (which is the actual branch of the animal kingdom occupied by Flying Foxes) sounds way scarier than if I said it was the same weight as the Coppery Titi.
The fact the Fluffy Glider is ALSO one kilo didn’t help me one bit. But at last I could latch onto something:
the Eastern Gray Squirrel is 1 kilo! OKAY… now we are on to something.
And so an Eastern Gray Squirrel is the size of a half-grown kitten? Seriously? A full-grown cat is the size of 2 squirrels? That’s not very convincing is it!
The credibility of this list is compromised by the list of 481 adult creatures that weigh ZERO. Zero? Come on… at least put them in at 0.1, or 0.3 kilos. They can’t be ZERO!
Especially the Ornate Shrew! For realsies? … the crown alone is a quarter of a kilo!

LIST OF ADULT MAMMALS RECORDED AT ZERO KILOS
Geoffroy’s tamarin
Brazilian bare-faced tamarin
Silvery-brown bare-face tamarin
Variegated squirrel
Golden bandicoot
Numbat
Franklin’s ground squirrel
Saddlebacked tamarin
Mauritius flying fox
Variable flying fox
Yellow-toothed cavy
Gray four-eyed opossum
Cotton-top tamarin
Brown four-eyed opossum
Ryukyu flying fox
Guayaquil squirrel
Moluccan naked-backed fruit bat
Western gray squirrel
Pygmy rabbit
Pygmy loris
Squirrel glider
Barbary ground squirrel
Quaira spiny rat
Fork-marked lemur
Prevost’s squirrel
Texas pocket gopher
Greater dwarf lemur
Fat-tailed dwarf lemur
Brazilian spiny rat
Red-tailed squirrel
Checkered elephant shrew
Thick-tailed opossum
Cayenne spiny rat
Woolly opossum
Great jerboa
Philippine tree shrew
Rodriguez flying fox
Richardson’s ground squirrel
Weid’s black-tufted-ear marmoset
Long-eared desert hedgehog
Geoffroy’s marmoset
Brazilian guinea pig
Common yellow-toothed cavy
Bushy-tailed woodrat
Mueller’s giant Sunda rat
Greater stick-nest rat
Hammer-headed fruit bat
Central American woolly opossum
Red-legged sun squirrel
Black four-eyed opossum
Striped possum
Lyle’s flying fox
Western needle-clawed galago
Horse-tailed squirrel
South African hedgehog
Black-pencilled marmoset
Straw-colored fruit bat
Little red flying fox
Norway rat
Eastern woodrat
Belding’s ground squirrel
Gundi
Great gerbil
Andean spiny rat
Peters’s climbing rat
White-naped weasel
Armored spiny rat
Cascade golden-mantled ground squirrel
Dwarf mongoose
Allen’s squirrel galago
Pallas’s squirrel
Fire-footed rope squirrel
Yellow-faced pocket gopher
Golden-backed tree rat
Chinchilla-rat
Silky anteater
Small pika
Calabar angwantibo
Gray-bellied squirrel
White-tufted-ear marmoset
Black-shouldered opossum
Giant mole-rat
Slender loris
Desert woodrat
Southern plains woodrat
Degus
Euphrates jerboa
Shark Bay Islands barred bandicoot
Greater hedgehog tenrec
East African mole rat
Thomas’s rope squirrel
European ground squirrel
Smith’s bush squirrel
Fat sand rat
Four-toed elephant shrew
White-throated woodrat
Red squirrel
Northern tree shrew
Little golden-mantled flying fox
Dassie rat
Rat
Tree shrew
Large tree shrew
Tuan
Senegal galago
Bolivian squirrel
Plantain squirrel
Deppe’s squirrel
Kintampo rope squirrel
Guianan squirrel
Hispid cotton rat
Shaw’s jird
Cape mole-rat
Streaked tenrec
Damaraland mole-rat
Lesser hedgehog tenrec
Speke’s gundi
Plains pocket gopher
Gray tree rat
South African galago
Three-striped ground squirrel
Brandt’s hamster
Thirteen-lined ground squirrel
Orange-bellied Himalayan squirrel
Round-tailed ground squirrel
Silvery mole-rat
Palestine mole rat
Chozchoz
Golden-mantled ground squirrel
Nelson’s antelope squirrel
Long-tailed weasel
Australian false vampire bat
Vinogradov’s jird
Northern flying squirrel
Northern palm squirrel
Vlei rat
Indian gerbil
Banner-tailed kangaroo rat
Spectral tarsier
Brush-tailed rabbit rat
Little ground squirrel
Ear-spot squirrel
Green bush squirrel
Greater Egyptian jerboa
African mole rat
Siberian flying squirrel
Bush rat
Collared pika
Leadbeater’s possum
Harris’s antelope squirrel
Asian garden dormouse
Western tarsier
Carpentarian rock rat
European water vole
Philippine tarsier
Siberian five-toed jerboa
Botta’s pocket gopher
Desert kangaroo rat
Australian swamp rat
Mulgara
Savanna gerbil
Ermine
Sugar glider
Nile kusu
Leschenault’s rousette
Congo rope squirrel
White-tailed mouse
Shorttailed opossum
White-tailed antelope squirrel
Golden hamster
Northern pocket gopher
Long-haired rousette
Central rock rat
Persian jird
Tuco-tuco
Indian palm squirrel
Mohave ground squirrel
North American pika
Libyan jird
Coruro
Eastern chipmunk
European mole, common mole
Ansell’s mole-rat
Eastern aquatic mole
Short bare-tailed opossum
Splendid climbing mouse
Long-eared chipmunk
Siberian chipmunk
Arctic lemming
Garden dormouse
Greater spear-nosed bat
Gambian epauletted fruit bat
Wahlberg’s epauletted fruit bat
Acacia rat
Red-tailed chipmunk
Robinson’s mouse opossum
Cape gerbil
Western pocket gopher
Malabar spiny dormouse
Greater short-nosed fruit bat
Townsend’s chipmunk
Slender squirrel
Bunny or coney rat
Smoky mouse
Speckled marsupial mouse
Norway lemming
Sand-colored rat
Colorado chipmunk
Greater bulldog bat
Low’s squirrel
Collared lemming
Thicket rat
Heermann’s kangaroo rat
Great fruit-eating bat
Gray mouse lemur
Lesser short-nosed fruit bat
Southern flying squirrel
Australian native mouse
Bushy-tailed jird
Heath mouse
Cliff chipmunk
Hairy-tailed mole
Cretan spiny mouse
Agile kangaroo rat
Long-tailed mouse
Woodland mouse
Lesser tree shrew
Five-toed jerboa
African water rat
Pyrenean desman
Rufous elephant shrew
Tundra vole
Ord’s kangaroo rat
Long-tailed tree mouse
Brown mouse lemur
Star-nosed mole
Julia Creek dunnart
Richardson’s collared lemming
Cape short-eared gerbil
Lesser Egyptian jerboa
South African broad-headed mouse
Golden spiny mouse
Cinnamon antechinus
Mid-day jird
Marsupial mole
Mongolian jird
Long-eared elephant shrew
Vole
Mitchell’s hopping mouse
Slender gerbil
Feather-tailed possum
Four-striped grass mouse
Darwin’s leaf-eared mouse
Water vole
Mexican mouse opossum
Pale gerbil
Talazaci’s long-tailed tenrec
Flat-headed vole
Common rock rat
Vesper rat
Cape elephant shrew
Meadow vole
North African elephant shrew
Rock mouse
Yellow-pine chipmunk
Red-nosed mouse
Pouched mouse
Common weasel
House shrew
Field vole
Rock dormouse
Yellow-footed marsupial mouse
Mountain pygmy possum
Spiny mouse
Greater red musk shrew
Least chipmunk
Pink fairy armadillo
Short-nosed elephant shrew
Pen-tailed tree shrew
Pale spear-nosed bat
Red-tailed phascogale
Merriam’s kangaroo rat
Jamaican fruit-eating bat
California mouse
Swamp antechinus
Panamanian spiny pocket mouse
Typical striped grass mouse
Dusky antechinus
Prairie vole
Brandt’s vole
Guatemalan deer mouse
Dormouse possum
Short-eared elephant shrew
Fat-tailed gerbil
Greater false vampire bat
Northern hopping mouse
Guinea multimammate mouse
Woolley’s pseudantechinus
Dobson’s long-tailed tenrec
Silver mountain vole
Greater Egyptian gerbil
Long-tailed vole
Fresno kangaroo rat
Australian hopping mouse
European free-tailed bat
Cloudy short-tailed opossum
Sundevall’s jird
Naked mole-rat
Red-cheeked dunnart
Fawn antechinus
Vampire bat
Korean field mouse
Western heather vole
Northern grasshopper mouse
Fat-tailed marsupial mouse
Florida mouse
Grey red-backed vole
Long-tailed pygmy possum
Lesser Egyptian gerbil
Tullberg’s rock rat
Little red kaluta
Yellow-necked mouse
Pencil-tailed tree mouse
Mouse-eared bat
Sandstone marsupial mouse
Noctule
Brown antechinus
Common vole
Eastern heather vole
Southern bog lemming
Japanese dormouse
Hazel dormouse
Forest dormouse
Murine mouse opossum
Woodland vole
Barbary striped grass mouse
Fat mouse
Wood lemming
Fish-eating bat
Woodland jumping mouse
Cactus mouse
Pinyon mouse
Monito del monte
New Zealand lesser short-tailed bat
Hoary bat
Woodland dormouse
Kultarr
Dzhungarian hamster
Lesser mouse-eared bat
Mediterranean pine vole
Big brown bat
White-footed mouse
Old World wood and field mouse
Greater horseshoe bat
Golden mouse
Southern grasshopper mouse
Short-tailed shrew
Striped field mouse
Paraguayan slender mouse opossum
White-lined broad-nosed bat
Pallid bat
Western jumping mouse
House mouse
Deer mouse
Highland gerbil mouse
Rio Napo mouse opossum
Red-backed mouse
European pine vole
Afghan mouse-like hamster
African spiny mouse
Steppe lemming
Stripe-faced dunnart
Little yellow-shouldered bat
Great Basin pocket mouse
Mouse-like hamster
Long-tailed pocket mouse
Particolored bat
Baluchistan gerbil
San Diego pocket mouse
Serotine
Philippine pygmy fruit bat
Fawn-colored mouse
Ningbing pseudantechinus
Gray-bellied dunnart
Forest shrew
Meadow jumping mouse
Canyon mouse
Lesser noctule
Geoffroy’s tailless bat
Fat-tailed dunnart
Eurasian water shrew
Seba’s short-tailed bat
Elliot’s short-tailed shrew
Pond bat
Southern water shrew
Oldfield mouse
Nelson’s pocket mouse
Townsend’s big-eared bat
Brant’s climbing mouse
Pygmy gliding possum
Brazilian free-tailed bat
Dark kangaroo mouse
Northern bat
Kimberly mouse
White-toothed shrew
Little long-tailed dunnart
Water shrew
California leaf-nosed bat
Western broad-nosed bat
Gray big-eared bat
Western pebble-mound mouse
Arizona pocket mouse
Bicolored white-toothed shrew
Geoffroy’s bat
Western harvest mouse
Rafinesque’s big-eared bat
Pilbara ningaui
Alston’s brown mouse
Ryukyu mouse
Ooldea dunnart
Bechstein’s bat
Cave myotis
Silver-haired bat
Western barbastelle
Little brown bat
Crowned shrew
Pygmy planigale
Lesser hairy-footed dunnart
Smaller horseshoe bat
Evening bat
Pallas’s long-tongued bat
Nathusius’s pipistrelle
Honey possum
Plains pocket mouse
Wongai ningaui
Gray myotis
Southern free-tailed bat
Eurasian shrew
Merriam’s pocket mouse
Northern pygmy mouse
Brown big-eared bat
Silky pocket mouse
Natterer’s bat
Alpine shrew
Eastern pipistrelle
Smoky shrew
Greater sac-winged bat
Lesser shrew
Trowbridge’s shrew
Little pocket mouse
Indiana bat
Northern birch mouse
Arctic shrew
Gray climbing mouse
Mouse
Brandt’s bat
Fulvus roundleaf bat
Eurasian pygmy shrew
Western long-eared myotis
Long-legged myotis
Large-eared tenrec
Paucident planigale
Keen’s myotis
Vagrant shrew
Eastern small-footed myotis
Ornate shrew
Old World harvest mouse
Kuhl’s pipistrelle
Montane shrew
Fringed myotis
Yuma myotis
Laxmann’s shrew
Whiskered bat
Lesser horseshoe bat
Common pipistrelle
Least shrew
Narrow-nosed planigale
Long-tailed planigale
Cinereus shrew
Black myotis
White-toothed pygmy shrew
So, if under a kilo, is it microevolution?
4 Likes
That definition can also run into problems. Do offspring have to be fertile? If mating is only successful 10% of the time, are they still in the same kind? What if mating is only successful 1% of the time, or 0.001% of the time? Where is the cut off?
I almost feel like discussing the possibility that humans and chimps could produce offspring, but that gets icky fast so I will leave it with just this mention.
I am more interested in the biology than the theological implications. It’s not that the two can’t be reconciled, only that I tend to focus on just one side of the picture. I also hedge my claims when talking about science which leads to blurry lines and large grey areas instead of the black and white categories that some prefer. I have always found it interesting that people will call this the “Species Problem”. A lot of biologists just view it as the “Species Reality”. Life doesn’t like being put into strict categories, and you can find exceptions for almost every general statement dealing with biology.
The definition may run into problems; I certainly agree with you on that. But this is nothing new when Biblical interpretation and Scientific methodologies bump together.
Naturally, you can be as scientific as you like. All I have to do on my side of the transom is follow the guidelines of Genesis 1:24-25:
“And God said, Let the earth bring forth the living creature after his kind, cattle, and creeping thing, and beast of the earth after his kind: and it was so. And God made the beast of the earth after his kind, and cattle after their kind, and every thing that creepeth upon the earth after his kind…”
Even in the human population, there are couples that are not particularly compatible reproductive. The criteria raised by verses 24-25 seems to be: in the natural state, can a population recreate the next generation?
So… maybe your “they don’t” criteria has a role! If we ask “do they”, and they don’t, problem solved!
But the question remains, if Jim Baker raises a whole pride of Ligers… are you going to fall on your sword and refuse to acknowledge them as a KIND (or part of a KIND)?
I don’t see any need for that. I think Christian Sciences (whoo… that almost involved you!) will just need to become more aware of when they are making a Scientific statement vs. when they are making a Theological statement that uses scientific terms.
As i reflect on the long post I wrote last night… there was lots of discussion about “species this” and “species that”. The Bible doesn’t require any discussion about “species”. So… that should get you and even your Christian colleagues OFF THE HOOK!
But then the topic turns THEOLOGICAL … and a colleague has to speak of “kinds” - - I believe Dr. Mayr would be satisfied with a definition of KINDS that depends on reproductive compatibility… regardless of whether it is naturally occurring or humanly sustained.
This shouldn’t be the deal-breaker. The deal-breaker should be: Biblical Kinds, once created, have to respond to the environment just like any other living thing - - and that means a Kind will evolve with each new generation!
What if it’s still too complicated, though? What if the essential truth in the statement is the simple concept that living things look more or less like their parents? The concept of heritability is really enormously important, and something we maybe take for granted.
If you break a rock into pieces the pieces will not retain the shape of the original. But take a shoot of a tree and grow it, and you can get a whole new tree. When flies lay eggs, the larvae turn into flies and not, say, centipedes.
Without heritability, evolution would be impossible. That was Darwin’s great insight over Lamarkianism, where animals change their shapes just by habit or striving. But heritable genetic variation makes the whole system effective.
My mistake, so you’re not claiming this is macroevolution. Then is it microevolution?
It’s been discussed but can you venture definitions of evolution, micro- and macroevolution?